Отличия от гомологичных образований
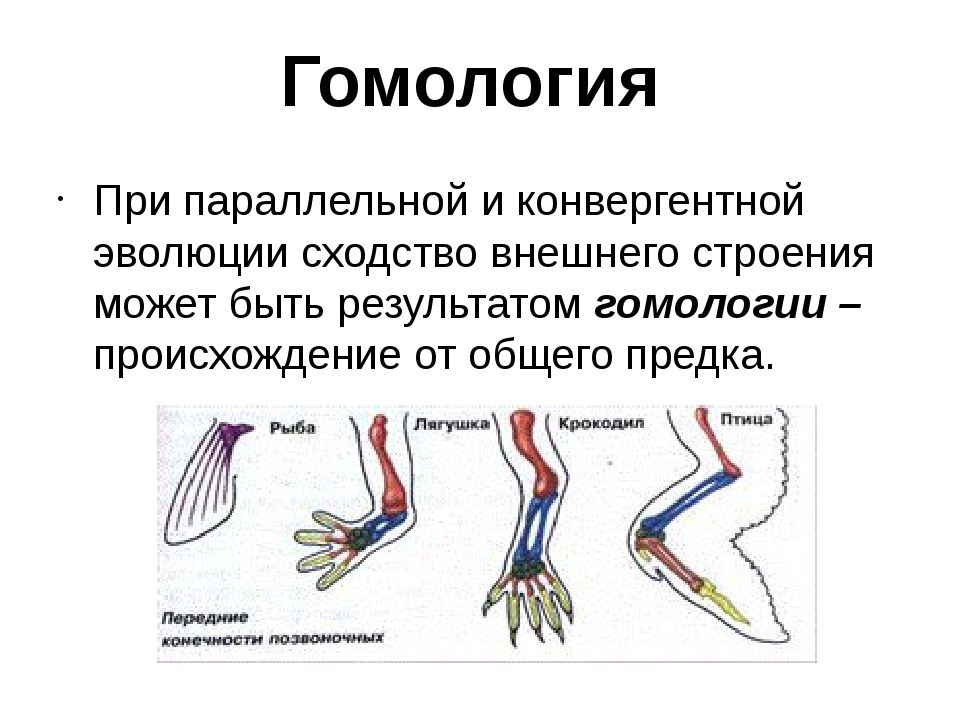
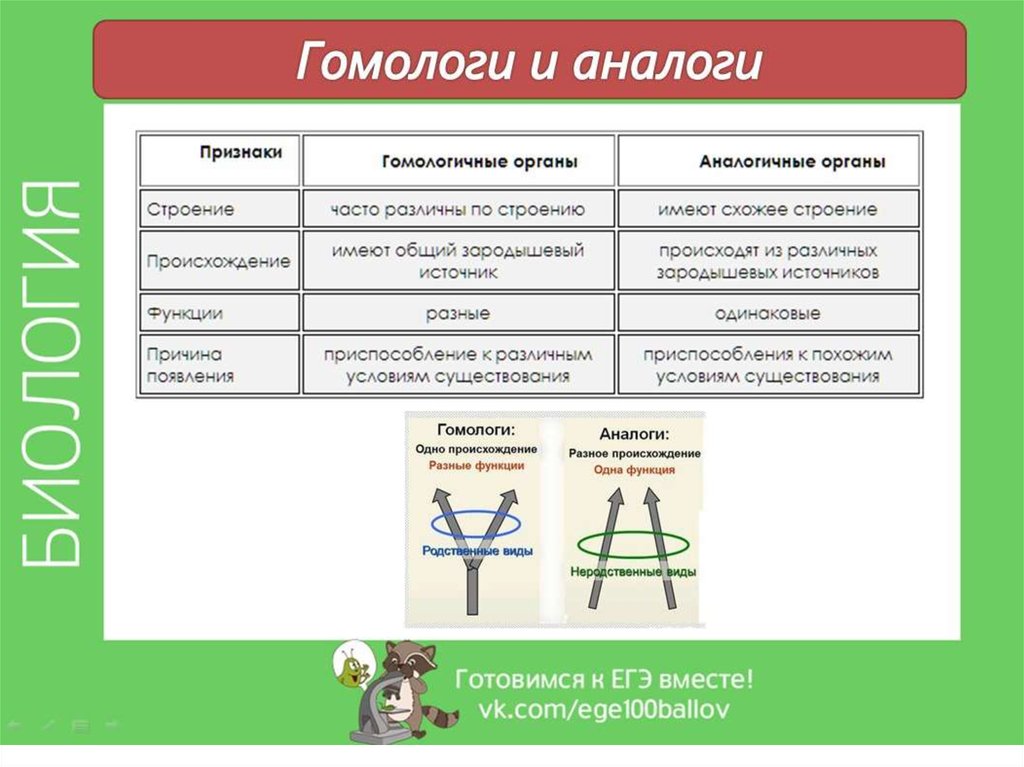
Гомологичные органы имеют общее происхождение. Какие органы называются аналогичными? Аналогичные структуры, напротив, происходят из совершенно разных частей организмов животных или растений. То есть, их зародышевые источники различны. Однако такие органы – результат приспособления к сходным условиям среды. Это отличает аналогичные органы от гомологичных, которые являются результатом приспособления к различным условиям. Внешне они иногда сильно отличаются у видов организмов.
Функции аналогичных органов всегда одинаковы. Виды, которые имеют такие сходные по функционированию органы, всегда не родственны друг другу.
Гомологичные и аналогичные органы
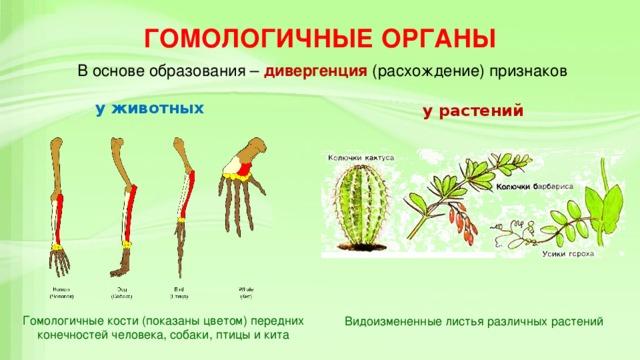
Некоторые органы животных и растений, несмотря на различные функции, имеют общее происхождение. К примеру, усики гороха прикрепляют растение к опоре, а шипы кактуса уменьшают интенсивность испарения воды. Но в обоих случаях эти структуры являются видоизменением листьев. Такое явление имеет свое название — гомология органов.
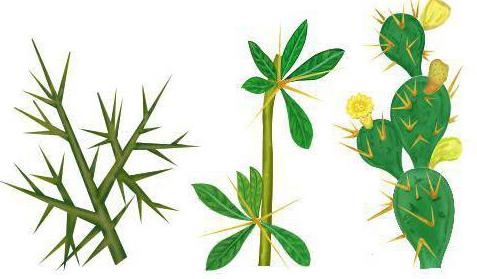
А вот иглы барбариса и шипы малины имеют разное происхождение. В первом случае это боковые листья, а во втором — производные покровной ткани растения. Такие органы называются аналогичными. Широкие крылья орла и бабочки также имеют разное происхождение. Хотя на первый взгляд определить это достаточно сложно, поскольку все эти структуры обеспечивают полет. Но у птиц это видоизмененные передние конечности, покрытые перьями. А у насекомых крылья представляют выросты покровов. Конечности же у них расположены под телом и не участвуют в полете.
Гомологичные и аналогичные органы являются прямым свидетельством общности происхождения различных животных. А различия в особенностях их строения обусловлены приспособлением к разным средам обитания и образу жизни.
История
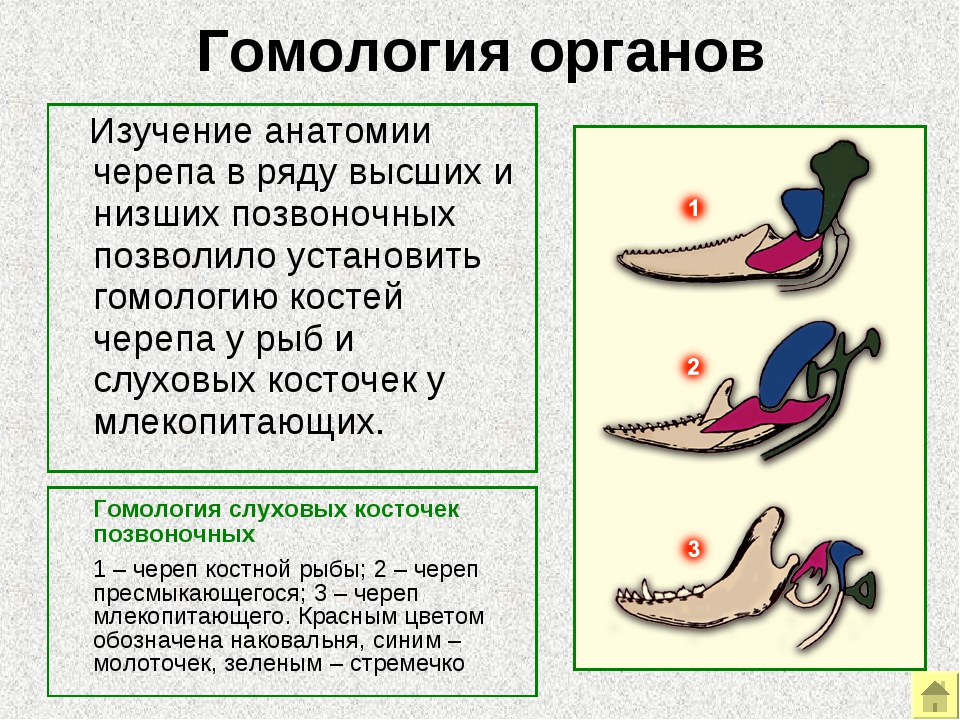
Понятие аналогии было введено Аристотелем и обозначало функциональное, и морфологическое сходство органов различных организмов. Ричард Оуэн уточнил это понятие как функциональное подобие, противоположное гомологии. Понятие гомологии в биологии было введено Ричардом Оуэном в 1840-е гг., не ставившего задачи решения филогенетических проблем. Он предложил различить аналогичные:
- «…a part or organ in one animal that has the same function as another part or organ in a different animal…»
и гомологичные структуры:
- «the same organ in different animals under every variety of form and function…»
Примерами аналогичных структур могут служить крылья насекомых и птиц. Примерами гомологичных — крыло птицы и рука человека.
Чарльз Дарвин (1859) считал, что аналогия возникает в ходе эволюции в сходных условиях жизни в результате приспособления к окружающей среде организмов далёких систематических групп (см. Конвергенция в биологии).
Гомология в сравнительной геномике
Гомологичные последовательности ДНК
Упрощенная схема эволюции глобинов. Каждый прямоугольник соответствует глобиновому гену. Узлы эволюционного дерева отмечены латинскими цифрами. Все глобины происходят от одного предшественника и, следовательно являются гомологами — ортологами протоглобина. Гемоглобины являются паралогами миоглобинов, так как произошли от гена протоглобина после его дупликации (на эволюционном отрезке между узлами I и II). Паралогами по отношению друг к другу являются, например, и гемоглобины человека: все они возникли в результате дупликаций и последующего накопления мутаций. Гемоглобины человека α1 и α2 являются ортологами α гемоглобинов акулы и курицы, так как происходят от про-α-гемоглобина общего предка, находящегося в узле II. То же верно и для β-гемоглобинов. При этом α-гемоглобины человека можно назвать паралогами, по отношению не только к человеческим, но и акульим, и куриным β-гемоглобинам, поскольку оба эти ряда ортологов восходят в конечном счете к одному протогемоглобину, возникшему на отрезке I—II.





Сравнительный анализ последовательностей нуклеотидов в ДНК и аминокислот в белках потребовал развития традиционного понятия гомологии. При анализе последовательностей принято различать ортологию и паралогию (и, соответственно, ортологи и паралоги).
Гомологичные последовательности называют ортологичными, если к их разделению привел акт видообразования: если ген существует у некоего вида, который дивергирует с образованием двух видов, то копии этого гена у дочерних видов называются ортологами. Гомологичные последовательности называют паралогичными, если к их разделению привело удвоение гена: если в пределах одного организма в результате хромосомной мутации произошло удвоение гена, то его копии называют паралогами.
Ортологи обычно выполняют идентичные или сходные функции. Это не всегда справедливо в отношении паралогов. В виду отсутствия давления отбора на одну из копий гена, подвергшегося удвоению, эта копия получает возможность беспрепятственно мутировать далее, что может привести к возникновению новых функций.
Так, например, гены, кодирующие миоглобин и гемоглобин, обычно считаются древними паралогами. Сходным образом, известные гены гемоглобинов (α, β, γ и т. д.) — паралоги друг друга. В то время как каждый из этих генов служит той же самой основной функции транспорта кислорода, их функции уже несколько дивергировали: гемоглобин зародыша (фетальный гемоглобин с субъединичной структурой α2γ2) имеет большее сродство к кислороду, чем гемоглобин взрослого человека (α2β2).
Другой пример: гены инсулина у крыс и мышей. У грызунов имеется пара паралогичных генов, однако вопрос о том, произошла ли дивергенция функций, остается открытым. Паралогичными обычно называют гены, принадлежащие одному и тому же виду, однако это вовсе не необходимо. Например, паралогами можно считать гены гемоглобина человека и миоглобина шимпанзе.
Одним из методов, применяющихся в современной биоинформатике для исследования гомологичности белков с известными аминокислотными последовательностями является выравнивание белков, суть которого заключается в нахождении с помощью различных алгоритмов наиболее консервативных остатков в этих последовательностях, которые обычно являются ключевыми для выполнения одной или нескольких функций белка, исследовании доменной структуры данного белка с помощью поиска известных структурных мотивов и доменов в исследуемом белке. Также с помощью различных баз данных можно осуществить поиск гомолога данного белка в различных организмах, построить филогенетическое дерево различных белковых последовательностей и тому подобное.
Гомологичные хромосомы
Гомологичными хромосомами в диплоидной клетке называют парные хромосомы, каждая из которых досталась от одного из родителей. За исключением половых хромосом у представителей гетерогаметного пола, последовательности нуклеотидов в каждой из гомологичных хромосом имеют значительное сходство по всей длине. Это означает, что в типичном случае они содержат одни и те же гены в одинаковой последовательности. Половые хромосомы у гетерогаметного пола также имеют гомологичные участки (хотя они занимают лишь часть хромосомы). С точки зрения анализа последовательностей, половые хромосомы следует считать паралогичными.
Примечания
- Гомология // : / гл. ред. А. М. Прохоров. — 3-е изд. — М. : Советская энциклопедия, 1969—1978.
- ↑
- Канаев И. И. Очерки из истории сравнительной анатомии до Дарвина. Развитие проблемы морфологического типа в зоологии. М.-Л.: Изд-во АН СССР, 1963. 299 с.
- Бляхер Л. Я. Проблемы морфологии животных. Исторические очерки. М.: Наука, 1976. 359 с.
- Беклемишев В. Н. Методология систематики. М.: KMK Scientific Press Ltd., 1994. 250 с.
- Owen, Richard. Lectures on Invertebrate Animals. London, 1843
- Darwin, Ch. On the origin of species by means of natural selection, or the preservation of the favoured races in the struggle for life. London, 1859
- (недоступная ссылка). Дата обращения 6 октября 2009.
- Догель В. А. 1954. Олигомеризация гомологичных органов как один из главных путей эволюции животных. Л.: Изд-во ЛГУ. 368 с.
- Боркин Л. Я., Наумов А. Д., Подлипаев С. Ф. 1971. Значение полимеризации и олигомеризации в эволюции систем органов // Вестник Ленинградского университета. № 21. С.7−18.
- Подлипаев С. А., Наумов А. Д., Боркин Л. Я. 1974. К определению понятий полимеризации и олигомеризации // Журнал общей биологии. Т.35. № 1. С.100−113.
- Городков К. Б. 1985. Олигомеризация и эволюция систем морфологических структур // Зоологический журнал Т.64. № 3. С.325−335
- Koonin EV, Galperin MY. «Sequence — Evolution — Function: Computational Approaches in Comparative Genomics.» Boston: Kluwer Academic, 2003.
- Вавилов Н. И. Закон гомологических рядов в наследственной изменчивости. Саратов, 1920. 16 с.
Гомология в сравнительной геномике
Гомологичные последовательности ДНК
 Упрощенная схема эволюции глобинов. Каждый прямоугольник соответствует глобиновому гену. Узлы эволюционного дерева отмечены римскими цифрами. Все глобины происходят от одного предшественника и, следовательно являются гомологами — ортологами протоглобина. Гемоглобины являются паралогами миоглобинов, так как произошли от гена протоглобина после его дупликации (на эволюционном отрезке между узлами I и II). Паралогами по отношению друг к другу являются, например, и гемоглобины человека: все они возникли в результате дупликаций и последующего накопления мутаций. Гемоглобины человека α1 и α2 являются ортологами α гемоглобинов акулы и курицы, так как происходят от про-α-гемоглобина общего предка, находящегося в узле II. То же верно и для β-гемоглобинов. При этом α-гемоглобины человека можно назвать паралогами, по отношению не только к человеческим, но и акульим, и куриным β-гемоглобинам, поскольку оба эти ряда ортологов восходят в конечном счете к одному протогемоглобину, возникшему на отрезке I—II.
Упрощенная схема эволюции глобинов. Каждый прямоугольник соответствует глобиновому гену. Узлы эволюционного дерева отмечены римскими цифрами. Все глобины происходят от одного предшественника и, следовательно являются гомологами — ортологами протоглобина. Гемоглобины являются паралогами миоглобинов, так как произошли от гена протоглобина после его дупликации (на эволюционном отрезке между узлами I и II). Паралогами по отношению друг к другу являются, например, и гемоглобины человека: все они возникли в результате дупликаций и последующего накопления мутаций. Гемоглобины человека α1 и α2 являются ортологами α гемоглобинов акулы и курицы, так как происходят от про-α-гемоглобина общего предка, находящегося в узле II. То же верно и для β-гемоглобинов. При этом α-гемоглобины человека можно назвать паралогами, по отношению не только к человеческим, но и акульим, и куриным β-гемоглобинам, поскольку оба эти ряда ортологов восходят в конечном счете к одному протогемоглобину, возникшему на отрезке I—II.
Сравнительный анализ последовательностей нуклеотидов в ДНК и аминокислот в белках потребовал развития традиционного понятия гомологии. При анализе последовательностей принято различать ортологию и паралогию (и, соответственно, ортологи и паралоги).
Гомологичные последовательности называют ортологичными, если к их разделению привел акт видообразования: если ген существует у некоего вида, который дивергирует с образованием двух видов, то копии этого гена у дочерних видов называются ортологами. Гомологичные последовательности называют паралогичными, если к их разделению привело удвоение гена: если в пределах одного организма в результате хромосомной мутации произошло удвоение гена, то его копии называют паралогами.
Ортологи обычно выполняют идентичные или сходные функции. Это не всегда справедливо в отношении паралогов. Ввиду отсутствия давления отбора на одну из копий гена, подвергшегося удвоению, эта копия получает возможность беспрепятственно мутировать далее, что может привести к возникновению новых функций.
Так, например, гены, кодирующие миоглобин и гемоглобин, обычно считаются древними паралогами. Сходным образом, известные гены гемоглобинов (α, β, γ и т. д.) — паралоги друг друга. В то время как каждый из этих генов служит той же самой основной функции транспорта кислорода, их функции уже несколько дивергировали: гемоглобин зародыша (фетальный гемоглобин с субъединичной структурой α2γ2) имеет большее сродство к кислороду, чем гемоглобин взрослого человека (α2β2).


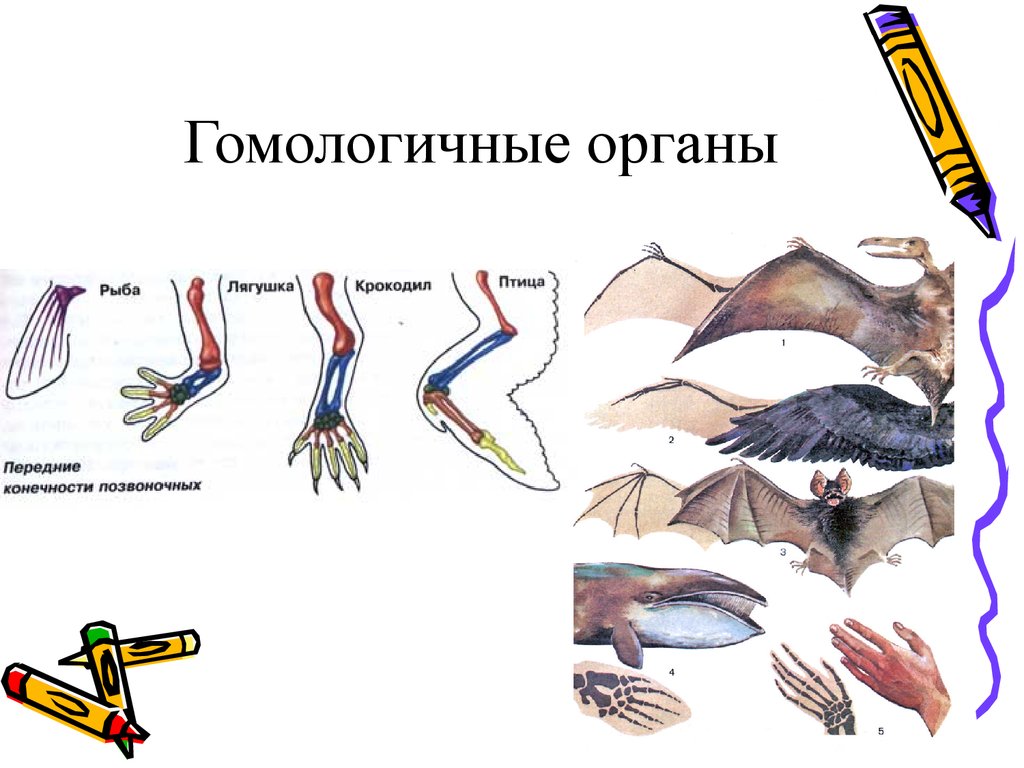

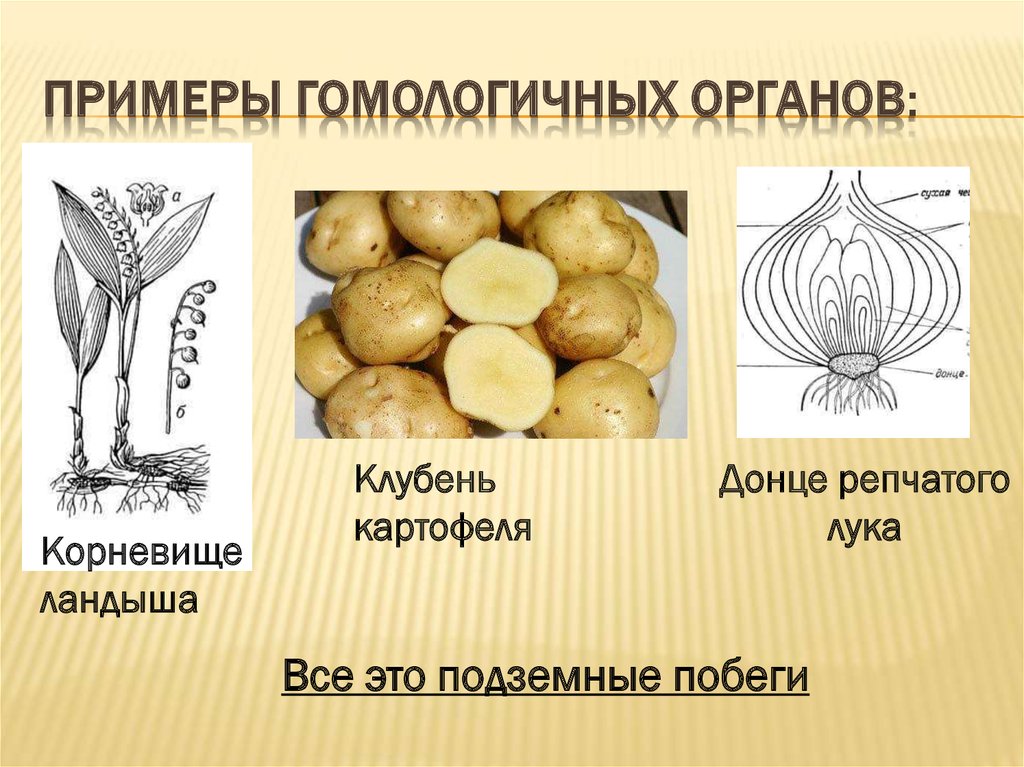
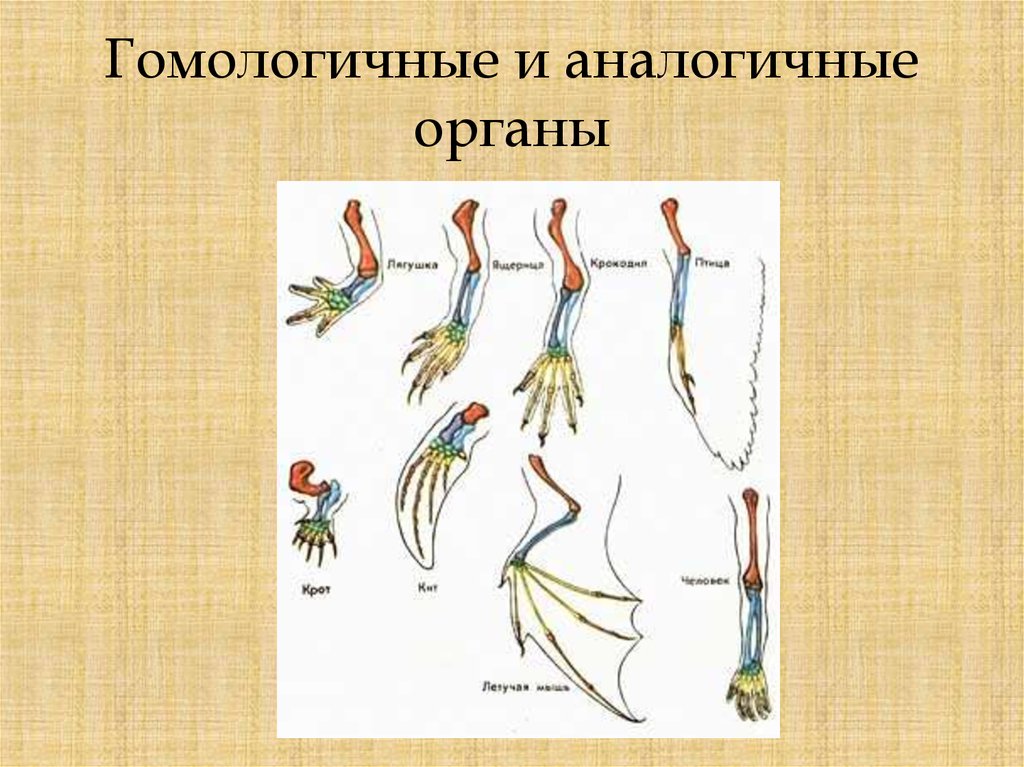

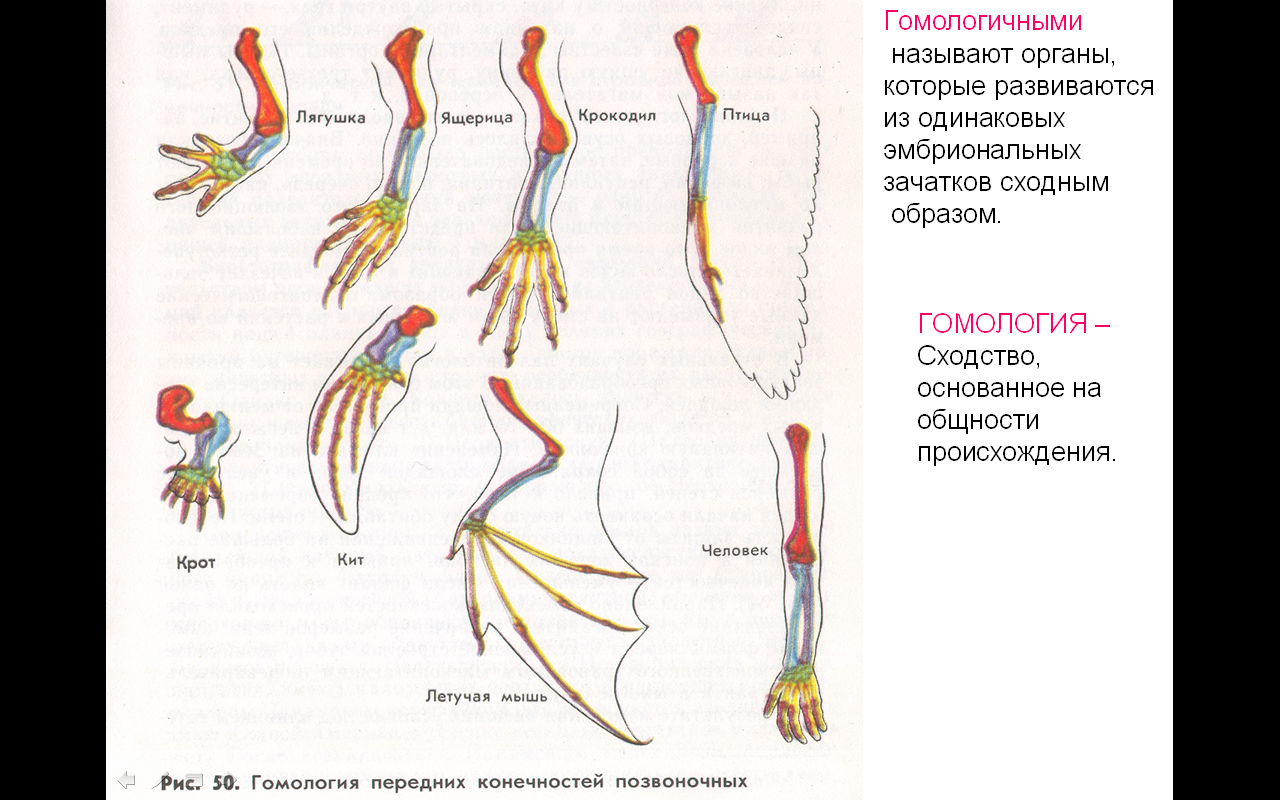

Другой пример: гены инсулина у крыс и мышей. У грызунов имеется пара паралогичных генов, однако вопрос о том, произошла ли дивергенция функций, остается открытым. Паралогичными обычно называют гены, принадлежащие одному и тому же виду, однако это вовсе не необходимо. Например, паралогами можно считать гены гемоглобина человека и миоглобина шимпанзе.
В современной биоинформатике для исследования гомологии белков с известными аминокислотными последовательностями применяется выравнивание белков, суть которого заключается в нахождении с помощью различных алгоритмов наиболее консервативных остатков в этих последовательностях, которые обычно являются ключевыми для выполнения одной или нескольких функций белка, исследовании доменной структуры данного белка с помощью поиска известных структурных мотивов и доменов в исследуемом белке. Также с помощью различных баз данных можно осуществить поиск гомолога данного белка в различных организмах, построить филогенетическое дерево различных белковых последовательностей и тому подобное.
Необходимо отметить, что иногда употребляемые термины «процент гомологии» и «значительная гомология» являются ошибочными, так как гомология последовательностей является понятием качественным, но не количественным. Гомологичные белки, например, могут сохранять лишь 10 % идентичных аминокислот, а негомологичные — иметь 30 % таковых.
Гомологичные хромосомы
Основная статья: Гомологичные хромосомы
Гомологичными хромосомами в диплоидной клетке называют парные хромосомы, каждая из которых досталась от одного из родителей. За исключением половых хромосом у представителей гетерогаметного пола, последовательности нуклеотидов в каждой из гомологичных хромосом имеют значительное сходство по всей длине. Это означает, что в типичном случае они содержат одни и те же гены в одинаковой последовательности. Половые хромосомы у гетерогаметного пола также имеют гомологичные участки (хотя они занимают лишь часть хромосомы). С точки зрения анализа последовательностей половые хромосомы следует считать паралогичными.
Олигомеризация гомологичных (гомодинамных) органов
Основная статья: Олигомеризация органов
Олигомеризация гомологичных (гомодинамных) органов — принцип Догеля — процесс (в ходе эволюции животных) уменьшения количества гомологичных и гомодинамных образований до некоторого определенного числа, связанный с интенсификацией функций системы. Реализуется в эволюции всех основных филогенетических стволов многоклеточных животных, сопровождаясь их прогрессивной морфологической и функциональной дифференцировкой.
Принцип множественной закладки новообразующихся органов Догеля — новые органы возникают (напр., из-за перемены образа жизни — перехода от сидячего образа жизни к подвижному или от водного к наземному) обычно в большом числе, слабо развиты, однородны и часто располагаются без определенного порядка. По мере дифференциации они приобретают определенную локализацию, количественно уменьшаясь до постоянного числа для данной таксономии. Например, сегментация тела в типе кольчатых червей носит множественный и неустановившийся характер. Все сегменты однородны. У членистоногих (произошедших от кольчатых червей) число сегментов в большинстве классов сокращается, становится постоянным, отдельные сегменты тела, объединяемые обычно в группы (голова, грудь, брюшко и т. п.), специализируются на выполнении определенных функций.
Выяснение, сохраняют они множественный характер или уже подверглись олигомеризации те или иные органы, позволяет судить о степени древности их возникновения. По комбинации органов разного возраста иногда можно судить о филогении.
Для эволюции одноклеточных характерна не олигомеризация, а полимеризация, то есть, увеличение, умножение частей клетки (органоидов).
Терминология
Сопоставление может быть как на уровне онтогенеза (индивидуального развития организма), так и на уровне филогенеза. В биологии понятие гомологии используется в сравнительной анатомии (см., например, Список гомологичных органов репродуктивной системы человека) с середины XIX века, и — в ревизованном виде — в сравнительных исследованиях генома. В рамках эволюционной биологии (уровень филогенеза) гомология интерпретируется как сходство, обусловленное происхождением от общего предшественника. В некотором смысле противоположным по значению термином, применяемым в тех случаях, когда два сходных органа или гена не имеют общего предшественника, является аналогия.
Кроме того, понятие гомологии используется в родственном, но несколько ином значении, в работах Н. И. Вавилова и более поздних авторов о законе гомологических рядов в наследственной изменчивости.








Гомологические ряды в наследственной изменчивости
В своей работе Закон гомологических рядов в наследственной изменчивостиНиколай Иванович Вавилов описал явления параллелизма мутаций в близкородственных группах растений. По аналогии с гомологическими рядами органических соединений, он предложил назвать это явление Гомологические ряды в наследственной изменчивости. Описание закономерностей наследственных вариаций позволяло предсказывать и целенаправленно искать ещё не выявленные гомологичные мутации у разных видов культурных растений, что привело к интенсификации селекционной работы.
Следует отметить, что, в отличие от химии, здесь речь идет об эмпирическом обобщении, а не о формальной теории, позволяющей выработать рациональную номенклатуру органических молекул, исходящую из определенного закона построения гомологического ряда.
Какие органы называются гомологичными: примеры
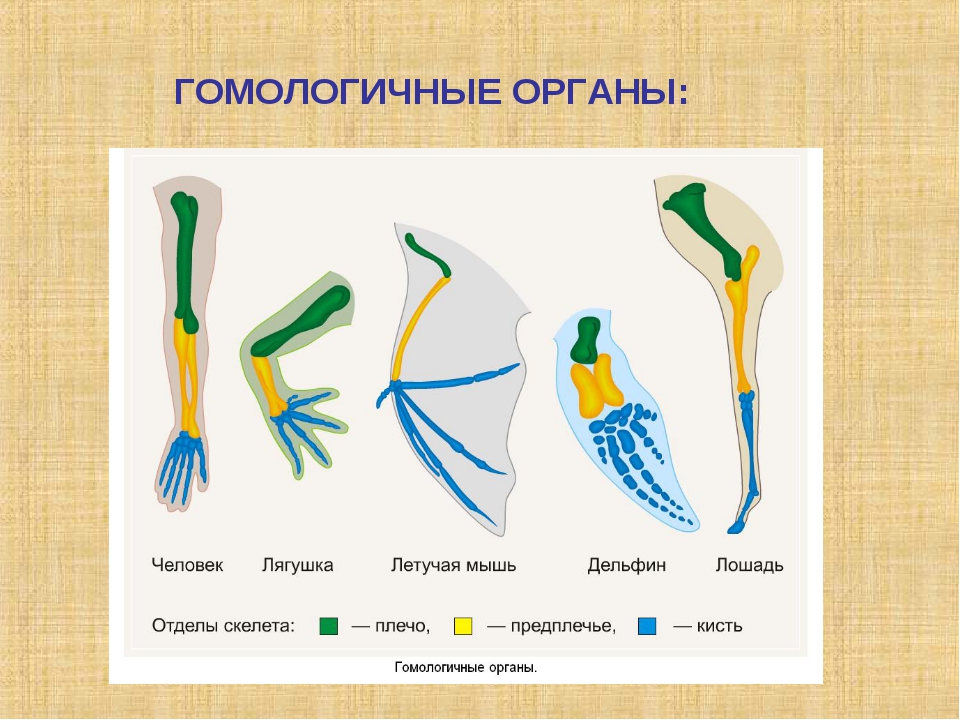
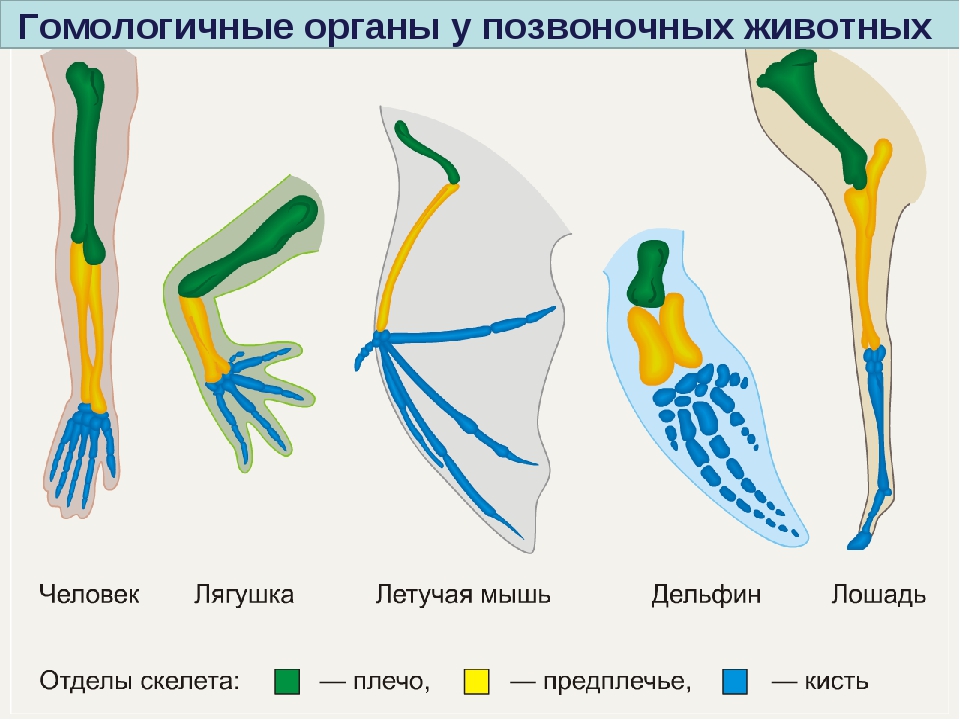
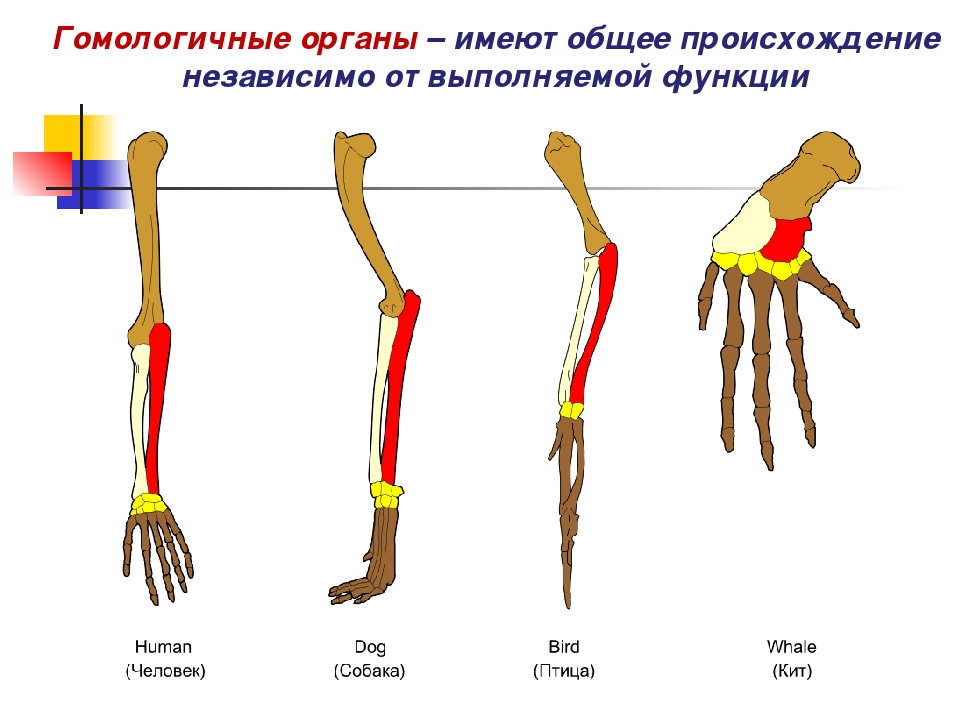
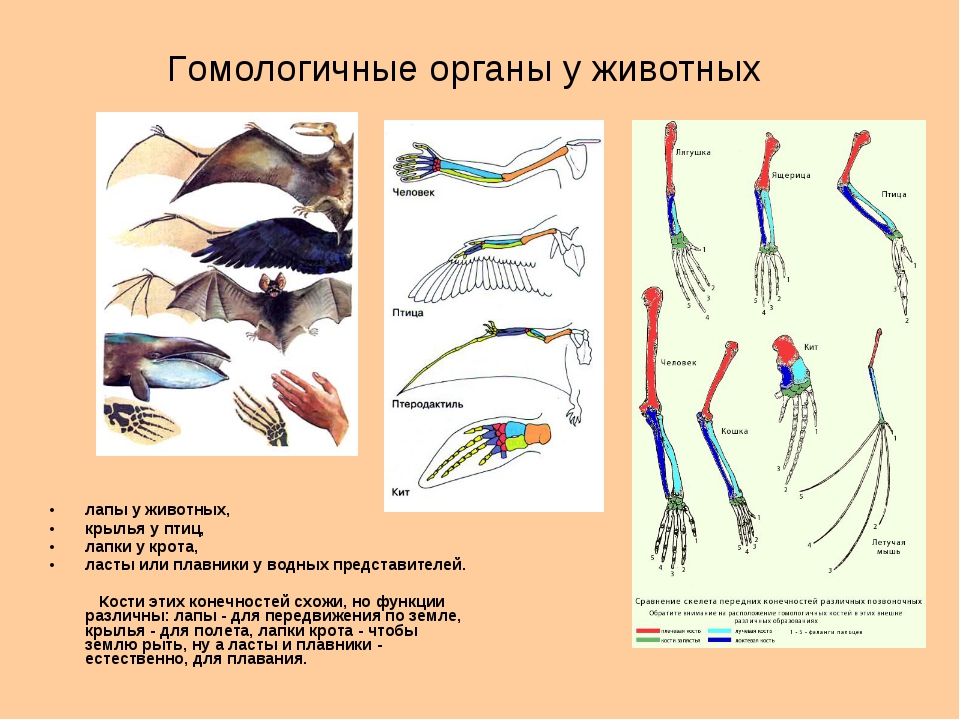
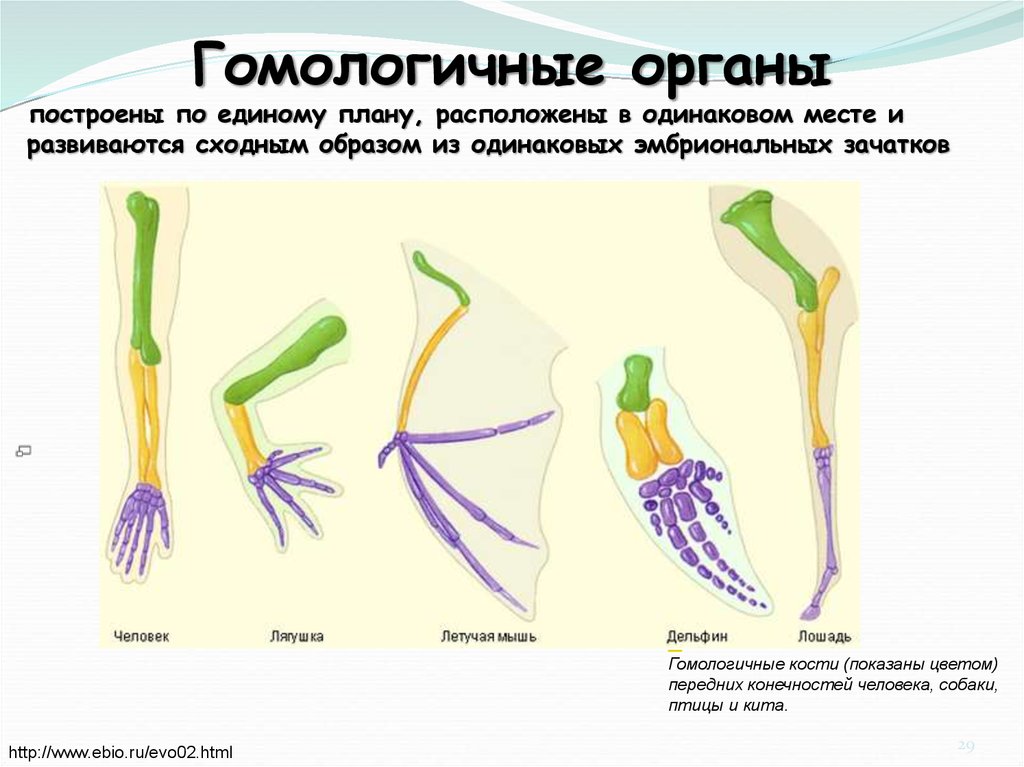
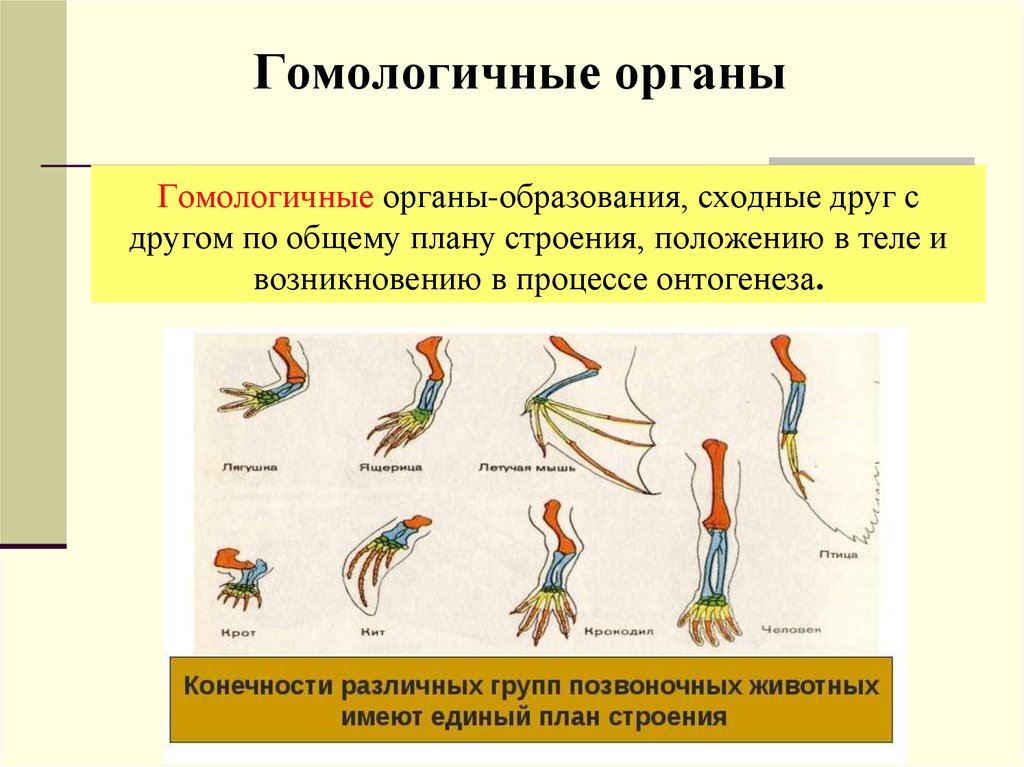
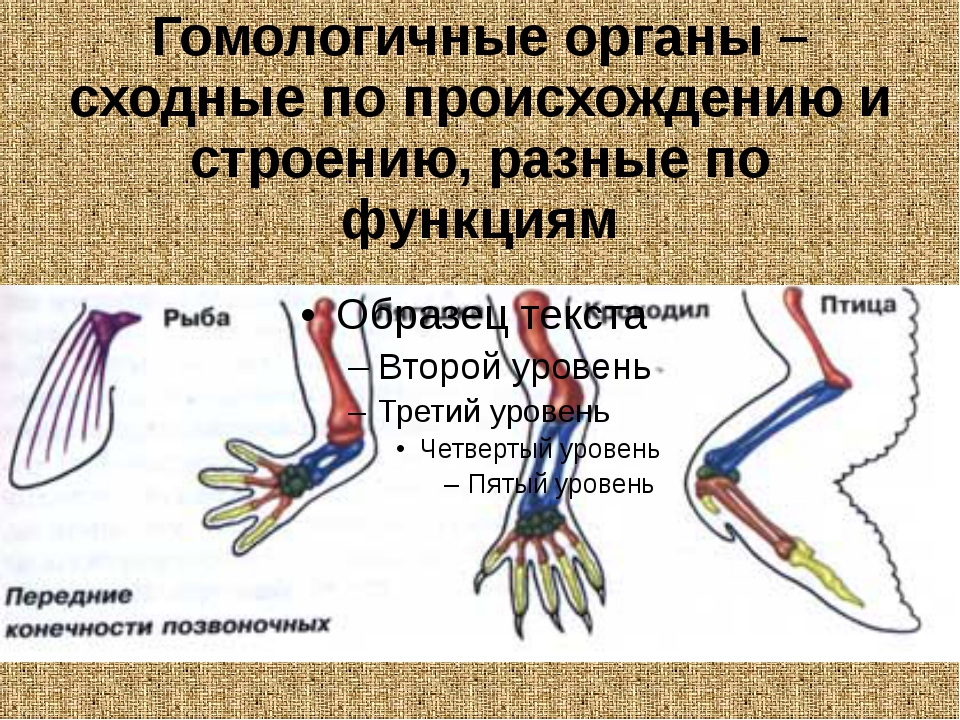
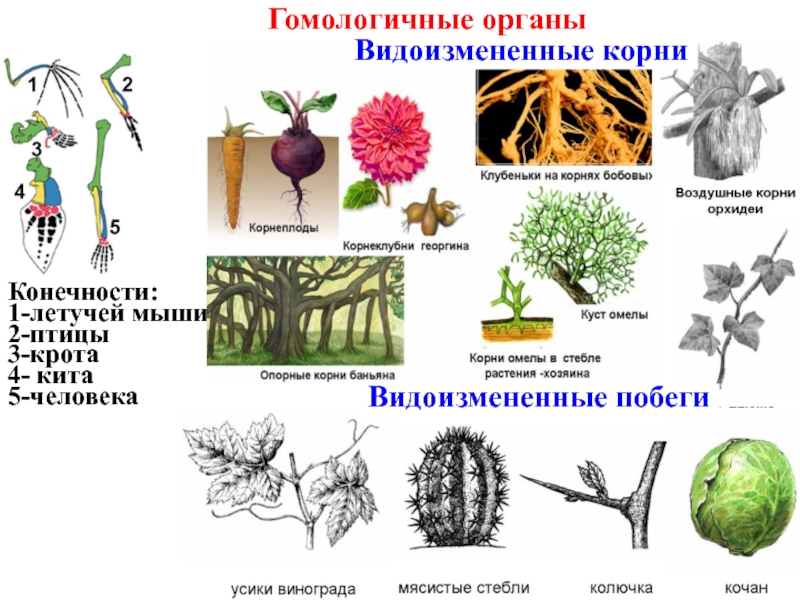
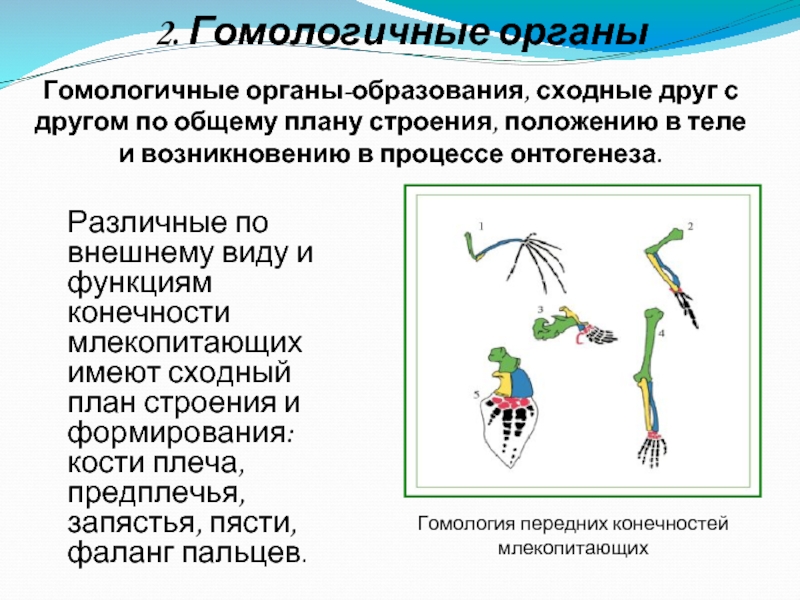
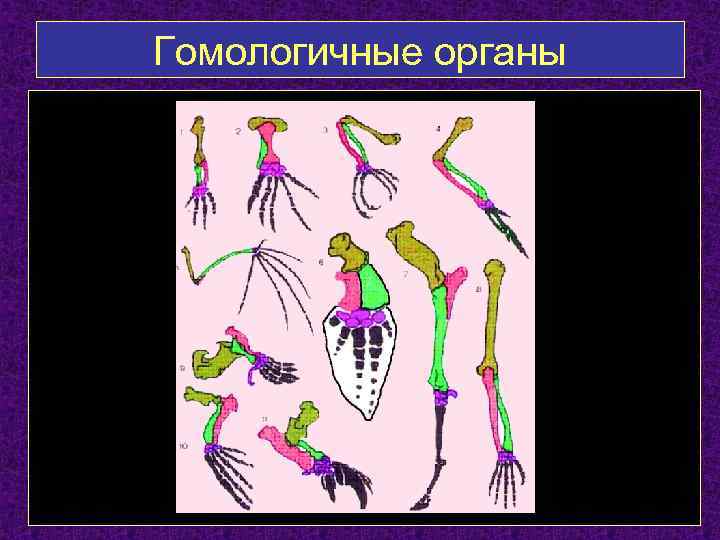
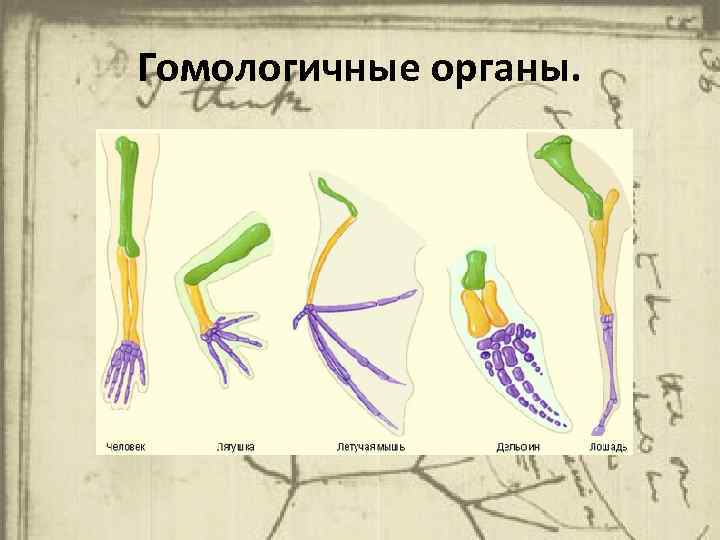
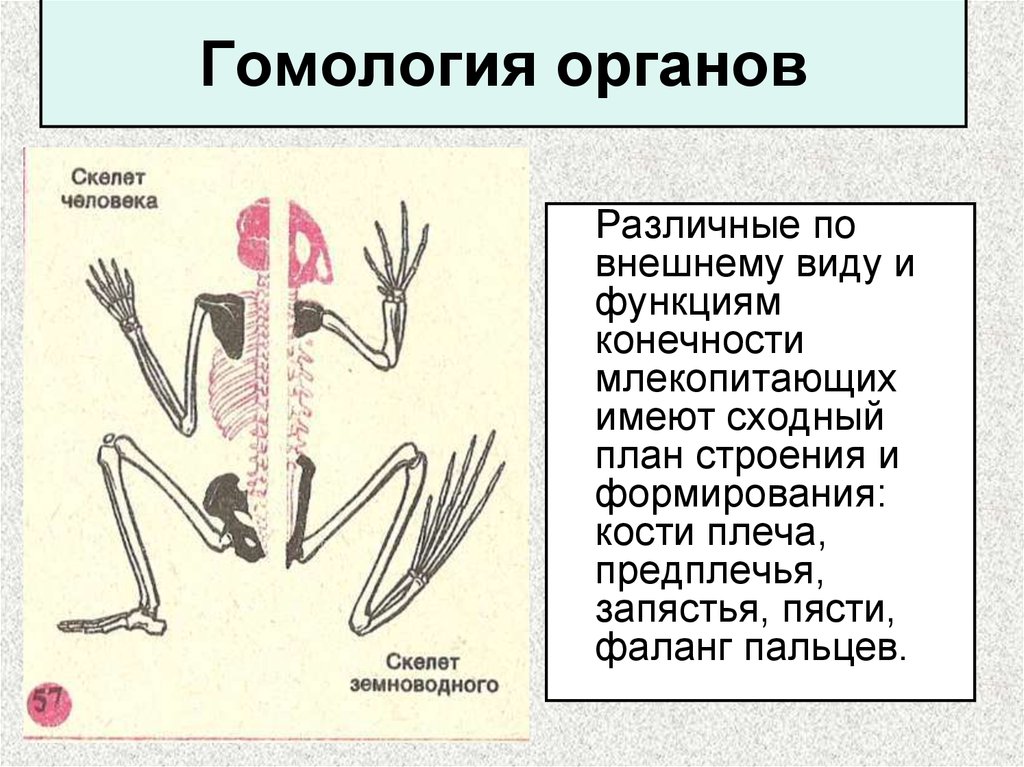
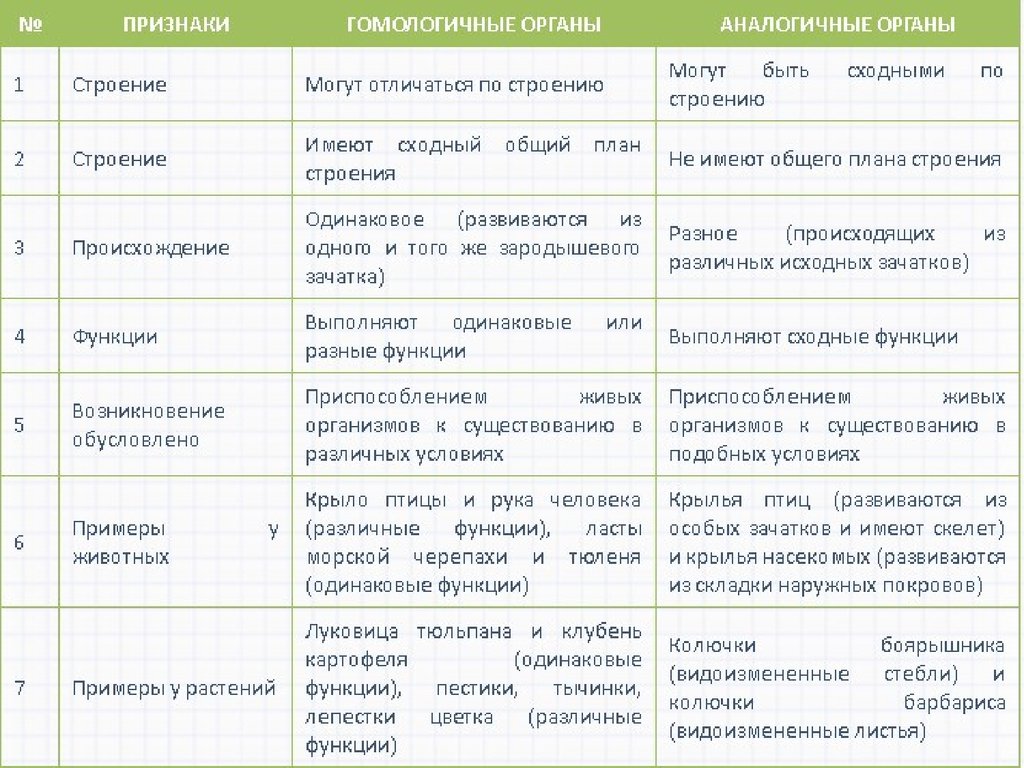
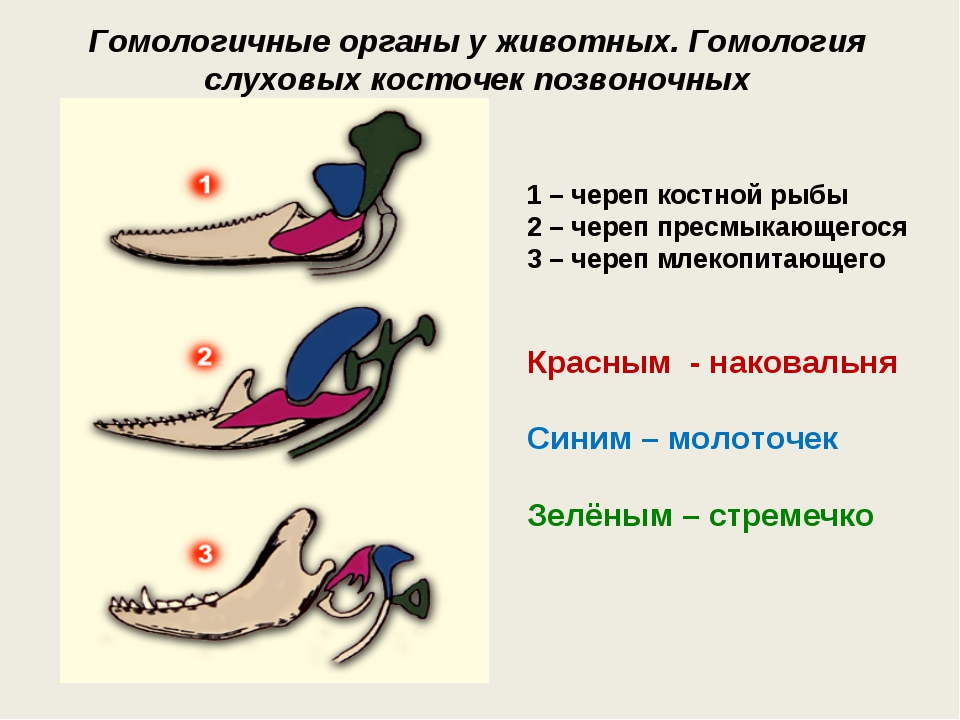
Самым типичным примером гомологий являются передние конечности позвоночных. Ласты кита и дельфина, крылья птицы и летучей мыши, руки человека, лапы крота и крокодила выполняют разные функции. Но строение их сходно. Все это передние конечности хордовых позвоночных, состоящие из трех отделов: плеча, предплечья и кисти.
К гомологичным органам относятся также видоизменения побегов различных растений. Они обладают значительными отличиями во внешнем строении и функциях. Корневище ландыша имеет удлиненные междоузлия, клубень картофеля накапливает запас воды с питательными веществами, а донце репчатого лука является основой для прикрепления мясистых листьев. Однако все гомологичные органы, примеры которых мы рассмотрели, имеют типичное для побега строение. Но и это еще не все!

видоизменений корней.

Ссылки
- Канаев И. И. Очерки из истории сравнительной анатомии до Дарвина. Развитие проблемы морфологического типа в зоологии. М.-Л.: Изд-во АН СССР, 1963. 299 с.
- Бляхер Л. Я. Проблемы морфологии животных. Исторические очерки. М.: Наука, 1976. 359 с.
- Беклемишев В. Н. Методология систематики. М.: KMK Scientific Press Ltd., 1994. 250 с.
- Owen, Richard. Lectures on Invertebrate Animals. London, 1843
- Darwin, Ch. On the origin of species by means of natural selection, or the preservation of the favoured races in the struggle for life. London, 1859
- Догель В. А. 1954. Олигомеризация гомологичных органов как один из главных путей эволюции животных. Л.: Изд-во ЛГУ. 368 с.
- Боркин Л. Я., Наумов А. Д., Подлипаев С. Ф. 1971. Значение полимеризации и олигомеризации в эволюции систем органов // Вестник Ленинградского университета. № 21. С.7−18.
- Подлипаев С. А., Наумов А. Д., Боркин Л. Я. 1974. К определению понятий полимеризации и олигомеризации // Журнал общей биологии. Т.35. № 1. С.100−113.
- Городков К. Б. 1985. Олигомеризация и эволюция систем морфологических структур // Зоологический журнал Т.64. № 3. С.325−335
- Вавилов Н. И. Закон гомологических рядов в наследственной изменчивости. Саратов, 1920. 16 с.
Примечания
- Гомология // : / гл. ред. А. М. Прохоров. — 3-е изд. — М. : Советская энциклопедия, 1969—1978.
- ↑
- Канаев И. И. Очерки из истории сравнительной анатомии до Дарвина. Развитие проблемы морфологического типа в зоологии. М.-Л.: Изд-во АН СССР, 1963. 299 с.
- Бляхер Л. Я. Проблемы морфологии животных. Исторические очерки. М.: Наука, 1976. 359 с.
- Беклемишев В. Н. Методология систематики. М.: KMK Scientific Press Ltd., 1994. 250 с.
- Owen, Richard. Lectures on Invertebrate Animals. London, 1843
- Darwin, Ch. On the origin of species by means of natural selection, or the preservation of the favoured races in the struggle for life. London, 1859
- (недоступная ссылка). Дата обращения 6 октября 2009.
- Догель В. А. 1954. Олигомеризация гомологичных органов как один из главных путей эволюции животных. Л.: Изд-во ЛГУ. 368 с.
- Боркин Л. Я., Наумов А. Д., Подлипаев С. Ф. 1971. Значение полимеризации и олигомеризации в эволюции систем органов // Вестник Ленинградского университета. № 21. С.7−18.
- Подлипаев С. А., Наумов А. Д., Боркин Л. Я. 1974. К определению понятий полимеризации и олигомеризации // Журнал общей биологии. Т.35. № 1. С.100−113.
- Городков К. Б. 1985. Олигомеризация и эволюция систем морфологических структур // Зоологический журнал Т.64. № 3. С.325−335
- Koonin EV, Galperin MY. «Sequence — Evolution — Function: Computational Approaches in Comparative Genomics.» Boston: Kluwer Academic, 2003.
- Вавилов Н. И. Закон гомологических рядов в наследственной изменчивости. Саратов, 1920. 16 с.
Другие примеры аналогичных органов
- Крылья птиц и крылья летавших в доисторическое время динозавров. Ученые выяснили, что современные птицы произошли от протоависа, а не от ящера–археоптерикса, как думали раньше. То есть, крылатые ящеры парят на крыльях, которые аналогичны крыльям птиц, но никак не гомологичны.
- Крылья птиц и крылья рукокрылых. Имеют разное происхождение. Летучие мыши тоже способны, как птицы, махать крыльями, а не только парить. Однако на гистологическом уровне строение органов, служащих для полета, чрезвычайно различно.
- Жабры рыб и жабры ракообразных. Оба таксона имеют органы, необходимые для дыхания в водной среде. Эти органы похожи тем, что усваивают кислород и потому имеют большую площадь поверхности. Для этого жабры образуют тонкие выросты. Органы дыхания рыб и ракообразных — пример аналогичных органов.
- Глаза позвоночных животных и насекомых. Органы зрения идентичны по функционированию. Строение тоже очень сходно: у простых глаз насекомых тоже имеется хрусталик, пигментные и светочувствительные клетки. Однако происхождение органов зрения насекомых не связано с происхождением глаз позвоночных животных. Сложные глаза насекомых — вершина эволюции органов зрения.
Гомологические ряды в наследственной изменчивости
В своей работе «Закон гомологических рядов в наследственной изменчивости»Николай Иванович Вавилов описал явления параллелизма мутаций в близкородственных группах растений. По аналогии с гомологическими рядами органических соединений он предложил назвать это явление Гомологические ряды в наследственной изменчивости. Описание закономерностей наследственных вариаций позволяло предсказывать и целенаправленно искать ещё не выявленные гомологичные мутации у разных видов культурных растений, что привело к интенсификации селекционной работы.
Следует отметить, что, в отличие от химии, здесь речь идет об эмпирическом обобщении, а не о формальной теории, позволяющей выработать рациональную номенклатуру органических молекул, исходящую из определенного закона построения гомологического ряда.
Примеры в царстве растений
- Колючки барбариса и боярышника. Острые образования барбариса – это видоизмененные листья. Колючки боярышника развиваются из побегов. Таким образом, острые выросты разных растений могут иметь различное происхождение, а, следовательно, быть аналогичными органами.
- Колючки кактуса – это видоизмененные листья, они только уменьшают площадь поверхности с устьицами, но и служат защитой от крупных травоядных животных. Шипы розы – это выросты кожицы, как и колючие образования шиповника – видоизмененные наросты коры. Шипы тоже защищают растения от животных.
- Корни аналогичны ризоидам. Оба вида органов служат для поглощения воды и питательных веществ из почвы. Однако корни – это сложно устроенный орган, тогда как ризоиды – это всего лишь выросты стебля. Ризоиды имеют мхи, лишайники, некоторые виды водорослей. Эти образования с первого взгляда не возможно отличить от настоящих корней.
- Семена аналогичны спорам. Оба образования служат для размножения и расселения растений. Семена устроены гораздо сложнее, имеют большой запас питательных веществ. Споры – легкие образования, разносятся ветром на значительные расстояния.
- Клубни растений могут происходить из стеблей либо из корней. Различие в происхождении говорит о наличии аналогии образований. Клубни стеблей и корней внешне очень похожи. Выполняют они одинаковую функцию: служат местом запаса питательных веществ.
Таким образом, аналогичные органы – это структуры организмов, сходные внешне и по внутреннему строению, а также выполняющие одинаковые функции. Однако такие структуры не происходят от общего первоначального образования.
Олигомеризация гомологичных (гомодинамных) органов
- Основная статья: Олигомеризация органов
Олигомеризация гомологичных (гомодинамных) органов — принцип Догеля — процесс (в ходе эволюции животных) уменьшения количества гомологичных и гомодинамных образований до некоторого определенного числа, связанный с интенсификацией функций системы.,,,. Реализуется в эволюции всех основных филогенетических стволов многоклеточных животных, сопровождаясь их прогрессивной морфологической и функциональной дифференцировкой.
Принцип множественной закладки новообразующихся органов Догеля — новые органы возникают (напр., из-за перемены образа жизни — перехода от сидячего образа жизни к подвижному или от водного к наземному) обычно в большом числе, слабо развиты, однородны и часто располагаются без определенного порядка. По мере дифференциации они приобретают определенную локализацию, количественно уменьшаясь до постоянного числа для данной таксономии. Например, сегментация тела в типе кольчатых червей носит множественный и неустановившийся характер. Все сегменты однородны. У членистоногих (произошедших от кольчатых червей) число сегментов в большинстве классов сокращается, становится постоянным, отдельные сегменты тела, объединяемые обычно в группы (голова, грудь, брюшко и т. п.), специализируются на выполнении определенных, функций.
Выяснение, сохраняют они множественный характер или уже подверглись олигомеризации те или иные органы, позволяет судить о степени древности их возникновения. По комбинации органов разного возраста иногда можно судить о филогении.




Для эволюции одноклеточных характерна не олигомеризация, а полимеризация, то есть, увеличение, умножение частей клетки (органоидов).
Олигомеризация гомологичных (гомодинамных) органов
Основная статья: Олигомеризация органов
Олигомеризация гомологичных (гомодинамных) органов — принцип Догеля — процесс (в ходе эволюции животных) уменьшения количества гомологичных и гомодинамных образований до некоторого определенного числа, связанный с интенсификацией функций системы. Реализуется в эволюции всех основных филогенетических стволов многоклеточных животных, сопровождаясь их прогрессивной морфологической и функциональной дифференцировкой.
Принцип множественной закладки новообразующихся органов Догеля — новые органы возникают (напр., из-за перемены образа жизни — перехода от сидячего образа жизни к подвижному или от водного к наземному) обычно в большом числе, слабо развиты, однородны и часто располагаются без определенного порядка. По мере дифференциации они приобретают определенную локализацию, количественно уменьшаясь до постоянного числа для данной таксономии. Например, сегментация тела в типе кольчатых червей носит множественный и неустановившийся характер. Все сегменты однородны. У членистоногих (произошедших от кольчатых червей) число сегментов в большинстве классов сокращается, становится постоянным, отдельные сегменты тела, объединяемые обычно в группы (голова, грудь, брюшко и т. п.), специализируются на выполнении определенных функций.
Выяснение, сохраняют они множественный характер или уже подверглись олигомеризации те или иные органы, позволяет судить о степени древности их возникновения. По комбинации органов разного возраста иногда можно судить о филогении.
Для эволюции одноклеточных характерна не олигомеризация, а полимеризация, то есть, увеличение, умножение частей клетки (органоидов).
Рудименты и атавизмы
Морфологическими доказательствами эволюции являются также рудиментарные органы. Это те части растений и животных, которые являются недоразвитыми. У человека это третье веко, второй ряд зубов, а также мышцы, которые двигают ушную раковину.
Признаками, противоположными рудиментам, являются атавизмы. Это проявление черт предков, не характерных для особей данного вида. В качестве примера можно привести развитие копчикового отдела позвоночника, многососковость, сплошной волосяной покров у человека. Если рассматривать животных, у них атавизмом является развитие задних конечностей у китов и змей.
Итак, гомологичные органы, примеры которых были рассмотрены в нашей статье, наряду с аналогиями, рудиментами и атавизмами являются морфологическими доказательствами процесса эволюции. Эти признаки проявляются как у животных, так и у растений. Гомологичными органами называют структуры, которые имеют общий план строения, но отличаются выполняемыми функциями. Наличие у человека перечисленных признаков доказывает его происхождение от животных в результате эволюционных преобразований.
Гомология в сравнительной геномике
Гомологичные последовательности ДНК
 Упрощенная схема эволюции глобинов. Каждый прямоугольник соответствует глобиновому гену. Узлы эволюционного дерева отмечены римскими цифрами. Все глобины происходят от одного предшественника и, следовательно являются гомологами — ортологами протоглобина. Гемоглобины являются паралогами миоглобинов, так как произошли от гена протоглобина после его дупликации (на эволюционном отрезке между узлами I и II). Паралогами по отношению друг к другу являются, например, и гемоглобины человека: все они возникли в результате дупликаций и последующего накопления мутаций. Гемоглобины человека α1 и α2 являются ортологами α гемоглобинов акулы и курицы, так как происходят от про-α-гемоглобина общего предка, находящегося в узле II. То же верно и для β-гемоглобинов. При этом α-гемоглобины человека можно назвать паралогами, по отношению не только к человеческим, но и акульим, и куриным β-гемоглобинам, поскольку оба эти ряда ортологов восходят в конечном счете к одному протогемоглобину, возникшему на отрезке I—II.
Упрощенная схема эволюции глобинов. Каждый прямоугольник соответствует глобиновому гену. Узлы эволюционного дерева отмечены римскими цифрами. Все глобины происходят от одного предшественника и, следовательно являются гомологами — ортологами протоглобина. Гемоглобины являются паралогами миоглобинов, так как произошли от гена протоглобина после его дупликации (на эволюционном отрезке между узлами I и II). Паралогами по отношению друг к другу являются, например, и гемоглобины человека: все они возникли в результате дупликаций и последующего накопления мутаций. Гемоглобины человека α1 и α2 являются ортологами α гемоглобинов акулы и курицы, так как происходят от про-α-гемоглобина общего предка, находящегося в узле II. То же верно и для β-гемоглобинов. При этом α-гемоглобины человека можно назвать паралогами, по отношению не только к человеческим, но и акульим, и куриным β-гемоглобинам, поскольку оба эти ряда ортологов восходят в конечном счете к одному протогемоглобину, возникшему на отрезке I—II.
Сравнительный анализ последовательностей нуклеотидов в ДНК и аминокислот в белках потребовал развития традиционного понятия гомологии. При анализе последовательностей принято различать ортологию и паралогию (и, соответственно, ортологи и паралоги).
Гомологичные последовательности называют ортологичными, если к их разделению привел акт видообразования: если ген существует у некоего вида, который дивергирует с образованием двух видов, то копии этого гена у дочерних видов называются ортологами. Гомологичные последовательности называют паралогичными, если к их разделению привело удвоение гена: если в пределах одного организма в результате хромосомной мутации произошло удвоение гена, то его копии называют паралогами.






Ортологи обычно выполняют идентичные или сходные функции. Это не всегда справедливо в отношении паралогов. Ввиду отсутствия давления отбора на одну из копий гена, подвергшегося удвоению, эта копия получает возможность беспрепятственно мутировать далее, что может привести к возникновению новых функций.
Так, например, гены, кодирующие миоглобин и гемоглобин, обычно считаются древними паралогами. Сходным образом, известные гены гемоглобинов (α, β, γ и т. д.) — паралоги друг друга. В то время как каждый из этих генов служит той же самой основной функции транспорта кислорода, их функции уже несколько дивергировали: гемоглобин зародыша (фетальный гемоглобин с субъединичной структурой α2γ2) имеет большее сродство к кислороду, чем гемоглобин взрослого человека (α2β2).
Другой пример: гены инсулина у крыс и мышей. У грызунов имеется пара паралогичных генов, однако вопрос о том, произошла ли дивергенция функций, остается открытым. Паралогичными обычно называют гены, принадлежащие одному и тому же виду, однако это вовсе не необходимо. Например, паралогами можно считать гены гемоглобина человека и миоглобина шимпанзе.
В современной биоинформатике для исследования гомологии белков с известными аминокислотными последовательностями применяется выравнивание белков, суть которого заключается в нахождении с помощью различных алгоритмов наиболее консервативных остатков в этих последовательностях, которые обычно являются ключевыми для выполнения одной или нескольких функций белка, исследовании доменной структуры данного белка с помощью поиска известных структурных мотивов и доменов в исследуемом белке. Также с помощью различных баз данных можно осуществить поиск гомолога данного белка в различных организмах, построить филогенетическое дерево различных белковых последовательностей и тому подобное.
Необходимо отметить, что иногда употребляемые термины «процент гомологии» и «значительная гомология» являются ошибочными, так как гомология последовательностей является понятием качественным, но не количественным. Гомологичные белки, например, могут сохранять лишь 10 % идентичных аминокислот, а негомологичные — иметь 30 % таковых.
Гомологичные хромосомы
Основная статья: Гомологичные хромосомы
Гомологичными хромосомами в диплоидной клетке называют парные хромосомы, каждая из которых досталась от одного из родителей. За исключением половых хромосом у представителей гетерогаметного пола, последовательности нуклеотидов в каждой из гомологичных хромосом имеют значительное сходство по всей длине. Это означает, что в типичном случае они содержат одни и те же гены в одинаковой последовательности. Половые хромосомы у гетерогаметного пола также имеют гомологичные участки (хотя они занимают лишь часть хромосомы). С точки зрения анализа последовательностей половые хромосомы следует считать паралогичными.