Дрожжеподобный таллом
Дрожжеподобный таллом – представлен отдельными клетками, покрытыми стенкой или распадающимися цепочками клеток (псевдомицелием), образующимся вследствие не расхождения материнских и дочерних клеток.
Дрожжевой таллом формируется в результате дегенерации мицелия. В ходе этого процесса каждый новый фрагмент мицелия отделяется септой от материнской клетки и обособляется, функционируя, как самостоятельный одноклеточный организм.
Дрожжевой таллом формируется обычно в жидких средах, богатых легкодоступными углеводами: в разлагающихся сочных плодах растений, гемолимфе насекомых, слизистых оболочках и мягких тканях позвоночных. При развитии в других условиях многие «дрожжи» способны формировать типичный мицелий.
Дрожжеподобный таллом характерен для многих представителей типа Аскомицеты (Ascomycota). Это так называемые «сумчатые дрожжи» – Sacharomyces, Schizosacharomyces. Такой же талломимеют некоторые виды типа Basidiomycota – «базидиальные дрожжи» – патогенные грибы Cryptococcus, Malassezia. Близок к дрожжевому таллом некоторых Zygomycota, в частности у патогенного для человека рода Basidiobolus.
Псевдомицелий – это разновидность дрожжевого таллома, образованная системой дрожжеподобных клеток, соединенных между собой клеточными стенками. При этом протопласты клеток полностью обособлены, но структурная взаимосвязь между клетками сохранена. Псевдомицелий в некоторых условиях формируется у Аскомицетов (род Candida) и Базидиомицетов (род Ustilago)
Слоевищные организмы
У многоклеточных водорослей таллом — это структура, которая может иметь нитчатую, объемную, пластинчатую или листовидную форму. Они растут исключительно в воде, поэтому не нуждаются ни в механических, ни в проводящих тканях. Их цвет зависит от вида пигмента, который преобладает в клетках. Однако независимо от этого признака все они содержат хлорофилл и способны к фотосинтезу. Большинство одноклеточных водорослей имеют таллом монадного типа. Он характеризуется постоянной формой и способностью к передвижению при помощи жгутиков. Реже встречаются низшие растения с амебоидным талломом. Благодаря тонкой оболочке они могут менять свою форму и передвигаться, образуя псевдоподии. Талломы монадного и амебоидного типа могут как быть одноклеточными, так и объединяться в колонии.







Тело грибов также образовано слоевищем. Оно состоит из отдельных нитей — гиф, совокупность которых называется мицелием или грибницей. Эти организмы питаются гетеротрофно, поглощая гифами воду с растворенными питательными веществами. Особый вид таллома имеют дрожжевые грибы. Они состоят из отдельных клеток или образованных ими цепочек, способных к почкованию.

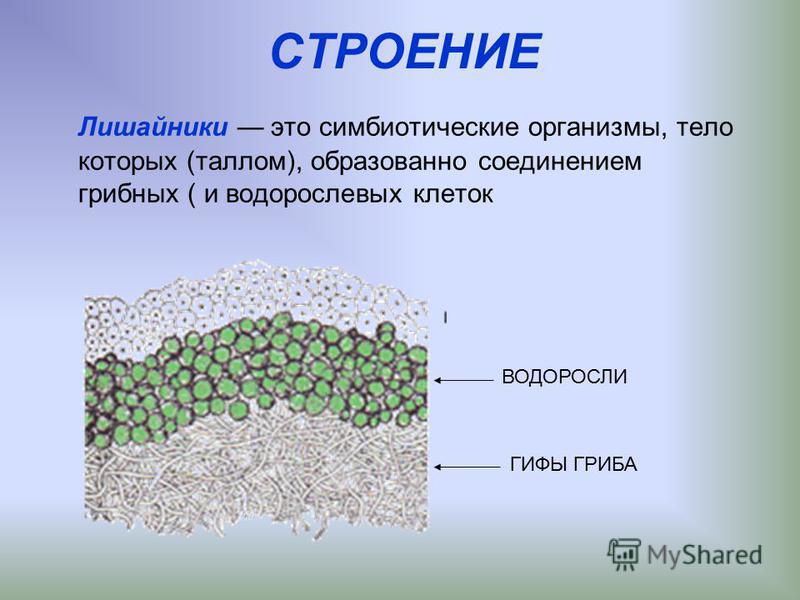
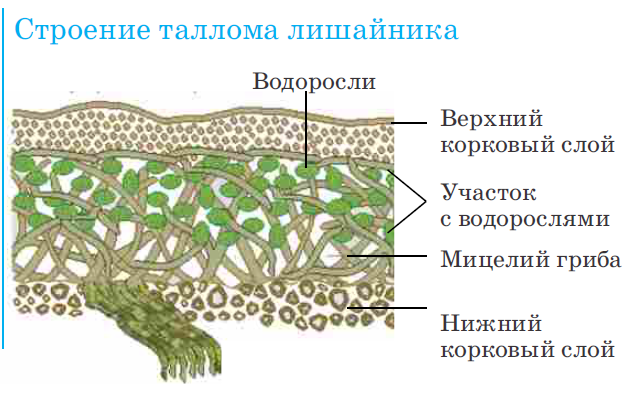
Талломы лишайников
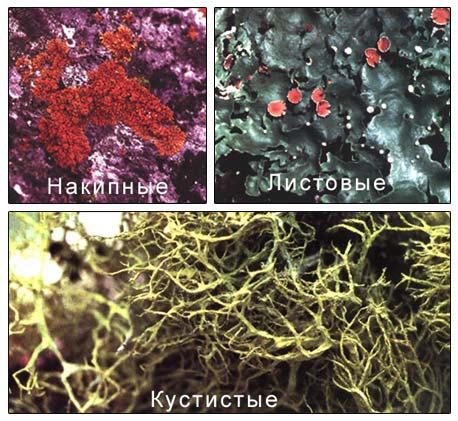
У лишайников по внешнему виду обычно выделяют талломы накипные, листоватые и кустистые.
- Накипной таллом характеризуются плагиотропным (горизонтальным) типом роста, имеют вид порошкообразного, зернистого налёта или корочки. Обычно такие лишайники прикрепляются к субстрату гифами сердцевинного слоя или подслоевища, нижний коровый слой, как правило, не сформирован. Накипные лишайники плотно срастаются с субстратом, порой отделить их от субстрата невозможно.
- Листоватые талломы также имеют плагиотропный тип роста, по форме напоминают более или менее рассечённые пластинки с хорошо сформированным нижним коровым слоем. Такие лишайники рыхло прикрепляются к субстрату с помощью специальных органов прикрепления, выростов нижнего корового слоя: ризин, гомфа или др.
- Кустистые талломы по уровню организации представляют собой высший этап развития таллома лишайника. В отличие от других они имеют ортотропный (вертикальный) тип роста гиф с верхушечным типом роста таллома. К субстрату они прикрепляются только небольшими участками нижней части таллома: тонкими нитевидными ризоидами, псевдогомфом и др. Внешне такие лишайники имеют вид кустиков — прямостоячих или повисающих.
Вертикальные выросты, которые образуются на горизонтальных слоевищах, называются подециями. Могут быть как простыми, так и сильно разветвлёнными. Верхняя часть подеция может иметь заличную форму — тупую, заострённую, сцифовидную (с кубковидным расширением). Подеции характерны для представителей родов Cladonia, Baeomyces, Stereocaulon.
Талломы лишайников
У лишайников по внешнему виду обычно выделяют талломы накипные, листоватые и кустистые.
- Накипной таллом характеризуются плагиотропным (горизонтальным) типом роста, имеют вид порошкообразного, зернистого налёта или корочки. Обычно такие лишайники прикрепляются к субстрату гифами сердцевинного слоя или подслоевища, нижний коровый слой, как правило, не сформирован. Накипные лишайники плотно срастаются с субстратом, порой отделить их от субстрата невозможно.
- Листоватые талломы также имеют плагиотропный тип роста, по форме напоминают более или менее рассечённые пластинки с хорошо сформированным нижним коровым слоем. Такие лишайники рыхло прикрепляются к субстрату с помощью специальных органов прикрепления, выростов нижнего корового слоя: ризин, гомфа или др.
- Кустистые талломы по уровню организации представляют собой высший этап развития таллома лишайника. В отличие от других они имеют ортотропный (вертикальный) тип роста гиф с верхушечным типом роста таллома. К субстрату они прикрепляются только небольшими участками нижней части таллома: тонкими нитевидными ризоидами, псевдогомфом и др. Внешне такие лишайники имеют вид кустиков — прямостоячих или повисающих.
Вертикальные выросты, которые образуются на горизонтальных слоевищах, называются подециями. Могут быть как простыми, так и сильно разветвлёнными. Верхняя часть подеция может иметь заличную форму — тупую, заострённую, сцифовидную (с кубковидным расширением). Подеции характерны для представителей родов Cladonia, Baeomyces, Stereocaulon.





Талломы грибов
У грибов и грибоподобных организмов (псевдогрибов) обычно выделяют талломы амёбоидные и плазмодиальные, мицелиальные и дрожжеподобные.
-
Амёбоидный таллом представляет собой одноядерную клетку, не имеющую клеточной стенки (имеется лишь цитоплазматическая мембрана и перипласт — уплотнённый наружный белковый слой). Плазмодиальный таллом имеет аналогичное строение, но содержит множество клеточных ядер. По морфологическим особенностям различают талломы:
- экстраматрикальные — находящиеся на поверхности и интраматрикальные — развивающиеся внутри субстрата;
- моноцентрические — представленные одной амёбоидной одноядерной клеткой, которая может иметь короткие безъядерные корнеподобные выросты (ризомицелий) и полицентрические — с удлинённым ризомицелием, образующим утолщения, содержащие дополнительные клеточные ядра;
- холокарпические — при развитии и переходе к стадии спороношения целиком превращающиеся в один спорангий и эукарпические — формирующие несколько спорангиев.
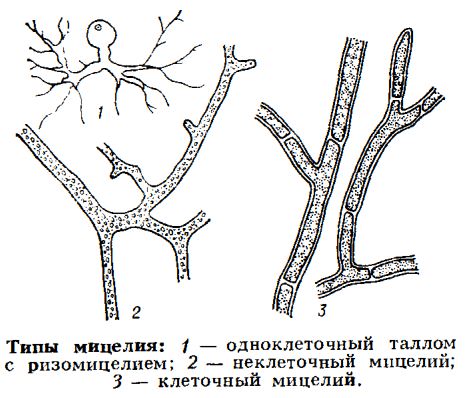
- Мицелиальный таллом состоит из тонких ветвящихся нитей — гиф, которые могут быть разделены септами на клетки (клеточный мицелий) или не иметь септ (неклеточный мицелий). Обычно гифы имеют неограниченный верхушечный рост, характерный как для первичной гифы, так и для боковых ветвей; гифы переплетаются и образуют анастомозы — слияния боковых ветвей; поэтому в зрелом мицелии обычно невозможно различить первичную гифу и ответвления. Из мицелия формируются почти все известные грибные структуры — плодовые тела, покоящиеся видоизменения (склероции, хламидоспоры), проводящие и поддерживающие структуры (синнемы, тяжи, ризоморфы), инфекционные структуры (аппрессории, гаустории, ловчие гифы).
- Дрожжеподобный таллом (см. Дрожжи) — отдельные клетки, имеющие клеточные стенки или короткие, легко распадающиеся цепочки клеток — псевдомицелий.
Особенности физиологии
Таллом, несмотря на достаточно примитивные черты строения, обеспечивает все необходимые процессы жизнедеятельности. С его помощью происходит вегетативное размножение и спорообразование. Клетки таллома являются хлорофиллоносными. Они способны поглощать готовые органические вещества из субстрата. Амебоидные и жгутиковые типы слоевищ способны к передвижению.
Талломы некоторых организмов могут вступать во взаимовыгодное сожительство с представителями других царств живой природы. К примеру, гифы грибов образуют микоризу с корнями высших растений. Составляющие таллома лишайника также получают выгоду от совместного существования. Гриб поглощает моносахариды, которые синтезирует клетка водоросли. Последние обеспечиваются минеральными веществами при помощи гифов.
Итак, таллом — это совокупность клеток, которые образуют тело низших и высших споровых растений, грибов и лишайников. Данная структура не образует тканей и состоит из неспециализированных структур, соединенных анатомически.
Что такое слоевище?
Форма большинства современных водорослей очень разнообразна, благодаря простому строению тела, которое в процессе эволюции изменялось и преобразовывалось, в зависимости от окружающей среды и других важных природных факторов. Тело организма, которое не разделено на отдельные функционально независимые органы и состоит из слабо специализированных тканей, — это слоевище. Это определение подходит не только для водорослей, но и для множества мхов, а также всех лишайников и некоторых грибов.
Слоевище является важным признаком простейшего развития растений и других организмов. Оно выполняет все функции сразу, которые принадлежали бы листьям, корню или стеблю по отдельности при более совершенной организации.







Так что такое слоевище? Это основная функциональная часть организма, которая отвечает за рост и размножение. Простота организации этой структуры в какой-то мере позволила освоить те места обитания, где не смогли бы произрастать современные виды растений. Наличие таллома можно назвать своеобразным плюсом.
Талломы водорослей
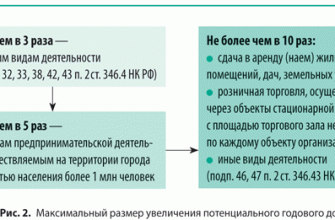
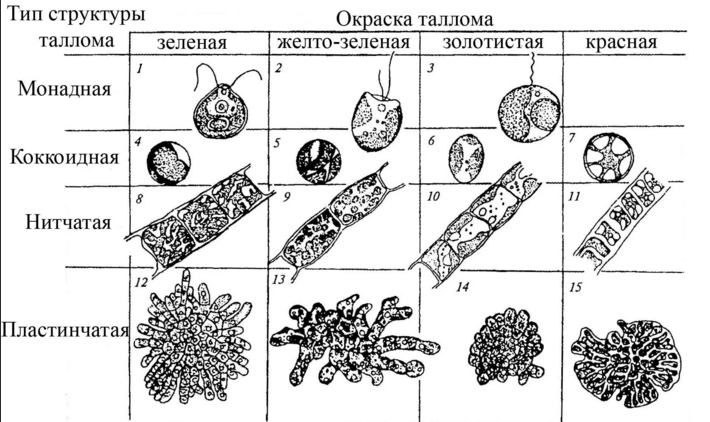
Талломы водорослей весьма разнообразны, в альгологии различают несколько типов морфологической дифференциации таллома. Часто, однако, чёткую границу между различными типами провести невозможно, так как существуют переходные формы.
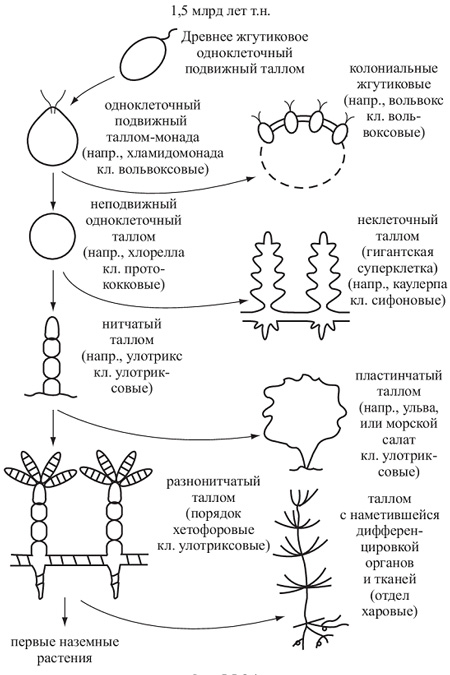
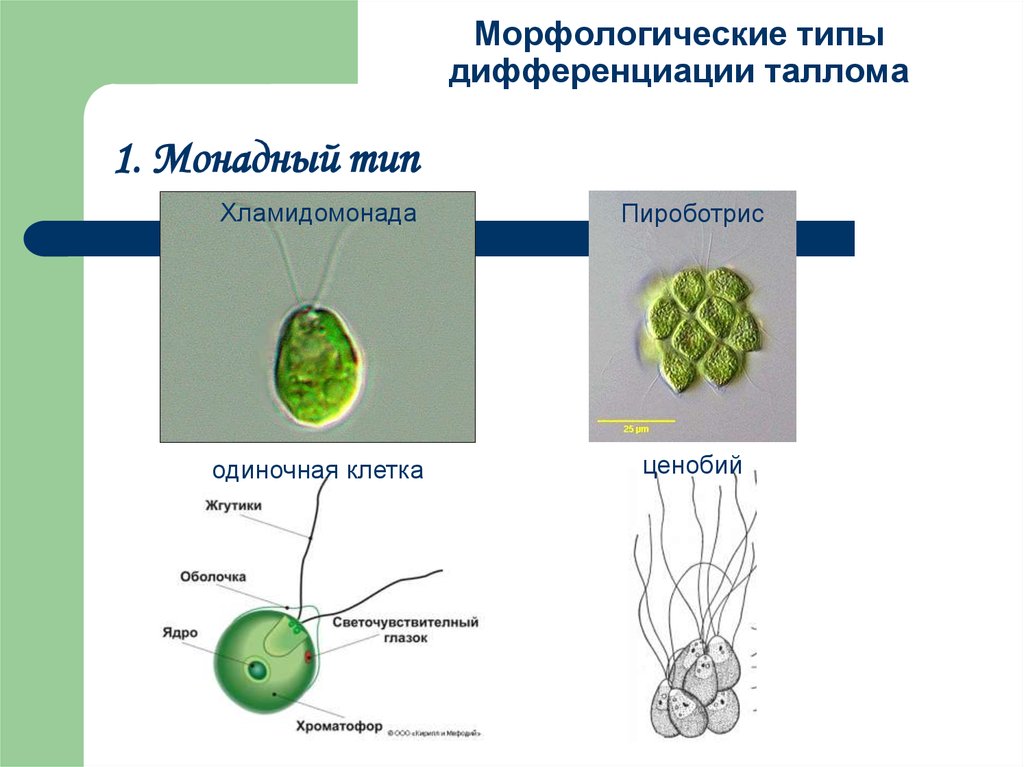
- Монадный, или жгутиковый тип — отдельные клетки, имеющие постоянную форму, способные к активному движению в водной среде при помощи жгутиков, одиночные или собирающиеся в колонии, ценобии (колонии с неизменным определённым числом клеток). Целые колонии и ценобии могут быть подвижными. У монадных водорослей часто встречаются характерные органеллы: центриоли, глоточный аппарат, пищеварительные вакуоли, стигма, сократительные вакуоли (у пресноводных). У многих отсутствуют вакуоли с клеточным соком и клеточная стенка, покровы монадных клеток разнообразны и отличаются от покровов клеток растений. Монадную структуру имеют также отдельные стадии водорослей (зооспоры, гаметы), обладающих многоклеточным талломом. Монадная организация таллома встречается у большинства таксонов водорослей, кроме хлорарахниофитовых, красных водорослей и цианобактерий.
- Амёбоидный, или ризоподиальный тип менее распространён, чем монадный. Амёбоидные клетки тоже могут быть одиночными или образовывать колонии, они лишены жёстких покровов, могут изменять свою форму и способны к амёбоидному движению. Встречается у хлорарахниофитовых, динофитовых и охрофитовых водорослей.
- Гемимонадный (пальмеллоидный, капсальный, трансгрессивный) тип — клетки, подобные монадным и имеющие характерные для монадного типа органеллы, но ведущие неподвижный образ жизни. Часто образуют колонии, состоящие из слизистых выделений и погружённых в эту слизь клеток. Встречается у зелёных, охрофитовых и глаукофитовых водорослей.
- Коккоидные клетки — одиночные или колониальные (ценобиальные), одетые в плотные оболочки, без жгутиков, в вегетативном состоянии неподвижные. Талломы коккоидного типа широко распространены у различных систематических групп водорослей, отсутствует этот тип у эвгленовых, гаптофитовых и криптофитовых.
- Сарциноидный тип является переходным между одноклеточными коккоидными и многоклеточными (паренхиматозными) водорослями. Клетки такого типа способны делиться в трёх взаимно перпендикулярных плоскостях и образовывать объёмные скопления. Как и для предыдущих типов, известны одиночные и колониальные представители сарциноидного типа. Насколько этот тип распространён в различных отделах водорослей, ещё окончательно не выяснено, характерные представители встречаются у зелёных водорослей, возможно также у класса жёлто-зелёных водорослей (отдел Охрофиты).
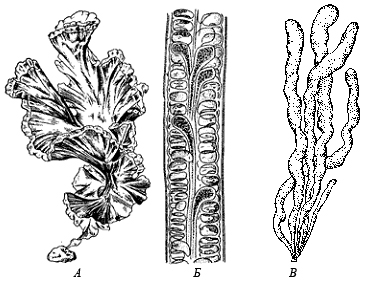
- Нитчатый, или трихальный тип — многоклеточный таллом, клетки которого делятся преимущественно в одной плоскости и образуют нити толщиной в один или несколько рядов. Нити могут быть простыми или разветвлёнными. Чаще всего клетки нити делятся поперечными перегородками, что обеспечивает постоянный рост нити в длину. Трихальный тип таллома широко представлен у охрофитовых, зелёных, красных водорослей и у цианобактерий.
- Разнонитчатый (гетеротрихальный) таллом схож с ничтатым, но различные участки его морфологически дифференцированы и приспособлены к выполнению различных функций, например, могут иметься две системы нитей — стелющиеся по субстрату и отходящие от них вертикально. Разнонитчатые талломы встречаются у охрофитовых, зелёных и красных водорослей.
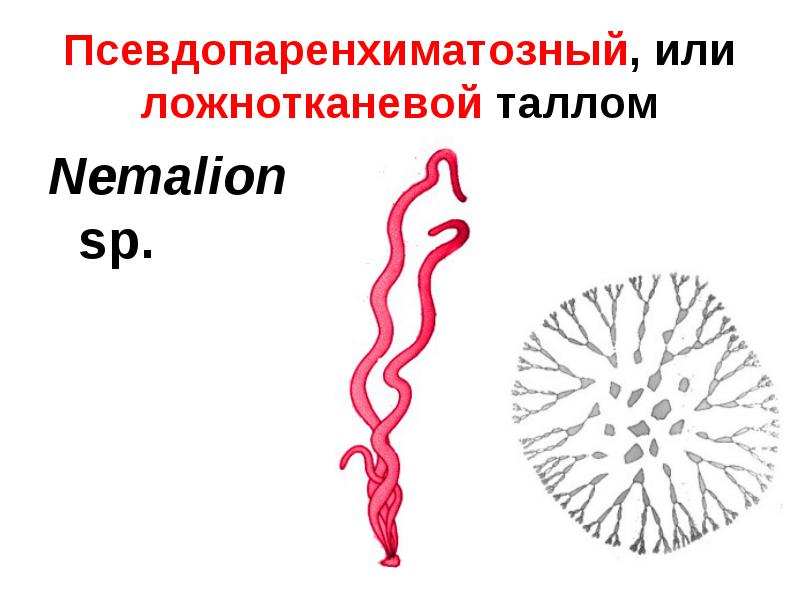
- Ложнотканевый, или псевдопаренхиматозный таллом представляет собой объёмистое слоевище, возникающее в результате срастания нитей. Встречается у охрофитовых и красных водорослей (возможно, также и у зелёных).
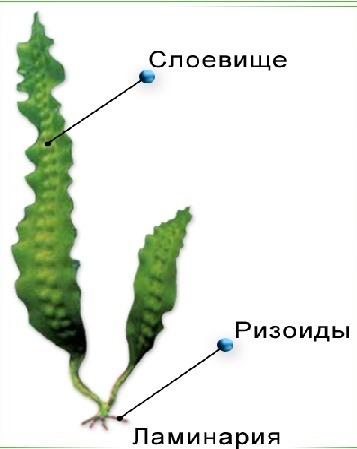
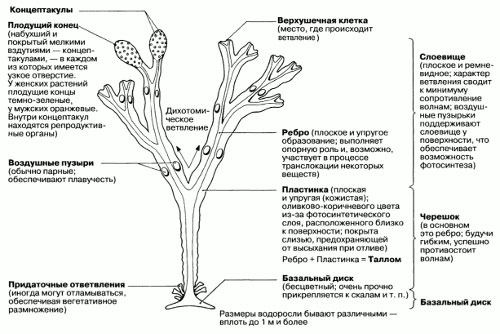
- Тканевый, пластинчатый, или паренхиматозный тип образуется за счёт деления клеток в двух или трёх направлениях. Образуются объёмные или пластинчатые, листовидные слоевища, дифференцированные на ткани, которые выполняют различные функции. Тканевые слоевища чаще всего встречаются у охрофитов (в частности, у бурых водорослей), известны они и у представителей зелёных и красных водорослей.
- Сифональный, или сифоновый тип — это многоядерное слоевище неклеточного строения. Оно может вырастать до макроскопических размеров и иметь определённую степень дифференцировки (внешнюю расчленённость). Перегородки появляются при повреждении такого таллома и в процессе размножения. Сифональные талломы известны у зелёных и жёлто-зелёных водорослей.
- Сифонокладальный таллом образуется из первичного сифонального и представляет собой сложное слоевище нитчатой или другой формы, состоящее из многоядерных элементов. Встречается только у зелёных водорослей.
Талломы водорослей
Талломы водорослей весьма разнообразны, в альгологии различают несколько типов морфологической дифференциации таллома. Часто, однако, чёткую границу между различными типами провести невозможно, так как существуют переходные формы.
- Монадный, или жгутиковый тип — отдельные клетки, имеющие постоянную форму, способные к активному движению в водной среде при помощи жгутиков, одиночные или собирающиеся в колонии, ценобии (колонии с неизменным определённым числом клеток). Целые колонии и ценобии могут быть подвижными. У монадных водорослей часто встречаются характерные органеллы: центриоли, глоточный аппарат, пищеварительные вакуоли, стигма, сократительные вакуоли (у пресноводных). У многих отсутствуют вакуоли с клеточным соком и клеточная стенка, покровы монадных клеток разнообразны и отличаются от покровов клеток растений. Монадную структуру имеют также отдельные стадии водорослей (зооспоры, гаметы), обладающих многоклеточным талломом. Монадная организация таллома встречается у большинства таксонов водорослей, кроме хлорарахниофитовых, красных водорослей и цианобактерий.
- Амёбоидный, или ризоподиальный тип менее распространён, чем монадный. Амёбоидные клетки тоже могут быть одиночными или образовывать колонии, они лишены жёстких покровов, могут изменять свою форму и способны к амёбоидному движению. Встречается у хлорарахниофитовых, динофитовых и охрофитовых водорослей.
- Гемимонадный (пальмеллоидный, капсальный, трансгрессивный) тип — клетки, подобные монадным и имеющие характерные для монадного типа органеллы, но ведущие неподвижный образ жизни. Часто образуют колонии, состоящие из слизистых выделений и погружённых в эту слизь клеток. Встречается у зелёных, охрофитовых и глаукофитовых водорослей.
- Коккоидные клетки — одиночные или колониальные (ценобиальные), одетые в плотные оболочки, без жгутиков, в вегетативном состоянии неподвижные. Талломы коккоидного типа широко распространены у различных систематических групп водорослей, отсутствует этот тип у эвгленовых, гаптофитовых и криптофитовых.
- Сарциноидный тип является переходным между одноклеточными коккоидными и многоклеточными (паренхиматозными) водорослями. Клетки такого типа способны делиться в трёх взаимно перпендикулярных плоскостях и образовывать объёмные скопления. Как и для предыдущих типов, известны одиночные и колониальные представители сарциноидного типа. Насколько этот тип распространён в различных отделах водорослей, ещё окончательно не выяснено, характерные представители встречаются у зелёных водорослей, возможно также у класса жёлто-зелёных водорослей (отдел Охрофиты).
- Нитчатый, или трихальный тип — многоклеточный таллом, клетки которого делятся преимущественно в одной плоскости и образуют нити толщиной в один или несколько рядов. Нити могут быть простыми или разветвлёнными. Чаще всего клетки нити делятся поперечными перегородками, что обеспечивает постоянный рост нити в длину. Трихальный тип таллома широко представлен у охрофитовых, зелёных, красных водорослей и у цианобактерий.
- Разнонитчатый (гетеротрихальный) таллом схож с ничтатым, но различные участки его морфологически дифференцированы и приспособлены к выполнению различных функций, например, могут иметься две системы нитей — стелющиеся по субстрату и отходящие от них вертикально. Разнонитчатые талломы встречаются у охрофитовых, зелёных и красных водорослей.
- Ложнотканевый, или псевдопаренхиматозный таллом представляет собой объёмистое слоевище, возникающее в результате срастания нитей. Встречается у охрофитовых и красных водорослей (возможно, также и у зелёных).
- Тканевый, пластинчатый, или паренхиматозный тип образуется за счёт деления клеток в двух или трёх направлениях. Образуются объёмные или пластинчатые, листовидные слоевища, дифференцированные на ткани, которые выполняют различные функции. Тканевые слоевища чаще всего встречаются у охрофитов (в частности, у бурых водорослей), известны они и у представителей зелёных и красных водорослей.
- Сифональный, или сифоновый тип — это многоядерное слоевище неклеточного строения. Оно может вырастать до макроскопических размеров и иметь определённую степень дифференцировки (внешнюю расчленённость). Перегородки появляются при повреждении такого таллома и в процессе размножения. Сифональные талломы известны у зелёных и жёлто-зелёных водорослей.
- Сифонокладальный таллом образуется из первичного сифонального и представляет собой сложное слоевище нитчатой или другой формы, состоящее из многоядерных элементов. Встречается только у зелёных водорослей.
Талломы грибов
У грибов и грибоподобных организмов (псевдогрибов) обычно выделяют талломы амёбоидные и плазмодиальные, мицелиальные и дрожжеподобные.
-
Амёбоидный таллом представляет собой одноядерную клетку, не имеющую клеточной стенки (имеется лишь цитоплазматическая мембрана и перипласт — уплотнённый наружный белковый слой). Плазмодиальный таллом имеет аналогичное строение, но содержит множество клеточных ядер. По морфологическим особенностям различают талломы:
- экстраматрикальные — находящиеся на поверхности и интраматрикальные — развивающиеся внутри субстрата;
- моноцентрические — представленные одной амёбоидной одноядерной клеткой, которая может иметь короткие безъядерные корнеподобные выросты (ризомицелий) и полицентрические — с удлинённым ризомицелием, образующим утолщения, содержащие дополнительные клеточные ядра;
- холокарпические — при развитии и переходе к стадии спороношения целиком превращающиеся в один спорангий и эукарпические — формирующие несколько спорангиев.
- Мицелиальный таллом состоит из тонких ветвящихся нитей — гиф, которые могут быть разделены септами на клетки (клеточный мицелий) или не иметь септ (неклеточный мицелий). Обычно гифы имеют неограниченный верхушечный рост, характерный как для первичной гифы, так и для боковых ветвей; гифы переплетаются и образуют анастомозы — слияния боковых ветвей; поэтому в зрелом мицелии обычно невозможно различить первичную гифу и ответвления. Из мицелия формируются почти все известные грибные структуры — плодовые тела, покоящиеся видоизменения (склероции, хламидоспоры), проводящие и поддерживающие структуры (синнемы, тяжи, ризоморфы), инфекционные структуры (аппрессории, гаустории, ловчие гифы).
- Дрожжеподобный таллом (см. Дрожжи) — отдельные клетки, имеющие клеточные стенки или короткие, легко распадающиеся цепочки клеток — псевдомицелий.
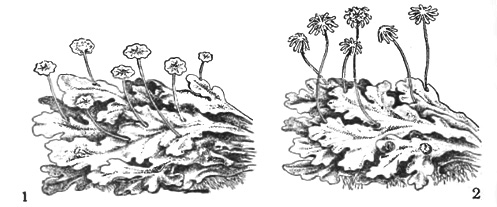
Талломы лишайников
У лишайников по внешнему виду обычно выделяют талломы накипные, листоватые и кустистые.
- Накипной таллом характеризуются плагиотропным (горизонтальным) типом роста, имеют вид порошкообразного, зернистого налёта или корочки. Обычно такие лишайники прикрепляются к субстрату гифами сердцевинного слоя или подслоевища, нижний коровый слой, как правило, не сформирован. Накипные лишайники плотно срастаются с субстратом, порой отделить их от субстрата невозможно.
- Листоватые талломы также имеют плагиотропный тип роста, по форме напоминают более или менее рассечённые пластинки с хорошо сформированным нижним коровым слоем. Такие лишайники рыхло прикрепляются к субстрату с помощью специальных органов прикрепления, выростов нижнего корового слоя: ризин, гомфа или др.
- Кустистые талломы по уровню организации представляют собой высший этап развития таллома лишайника. В отличие от других они имеют ортотропный (вертикальный) тип роста гиф с верхушечным типом роста таллома. К субстрату они прикрепляются только небольшими участками нижней части таллома: тонкими нитевидными ризоидами, псевдогомфом и др. Внешне такие лишайники имеют вид кустиков — прямостоячих или повисающих.
Вертикальные выросты, которые образуются на горизонтальных слоевищах, называются подециями. Могут быть как простыми, так и сильно разветвлёнными. Верхняя часть подеция может иметь заличную форму — тупую, заострённую, сцифовидную (с кубковидным расширением). Подеции характерны для представителей родов Cladonia, Baeomyces, Stereocaulon.





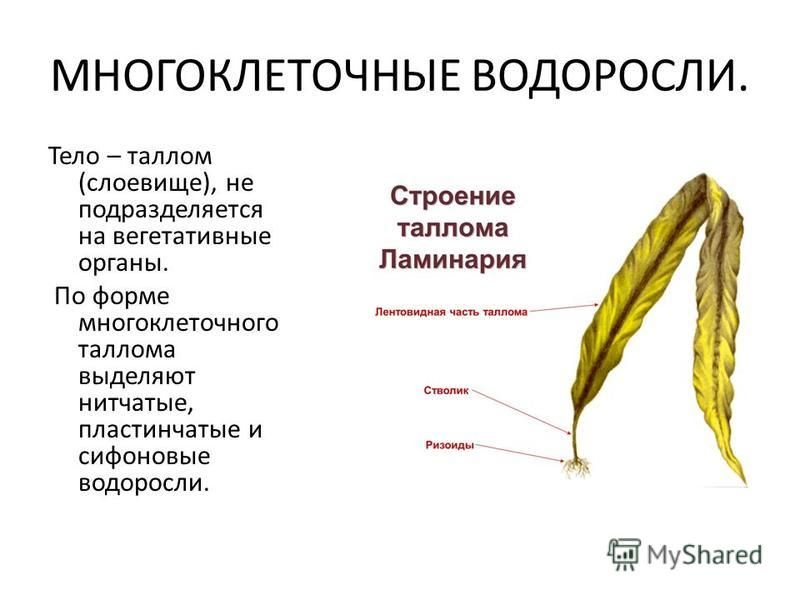

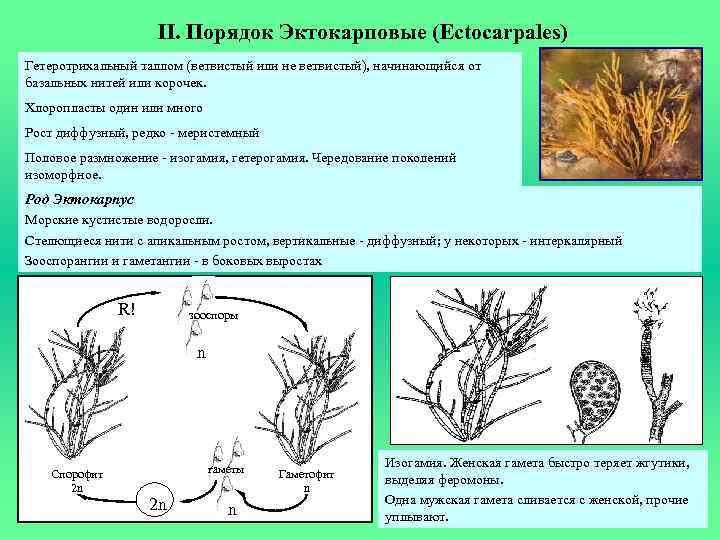
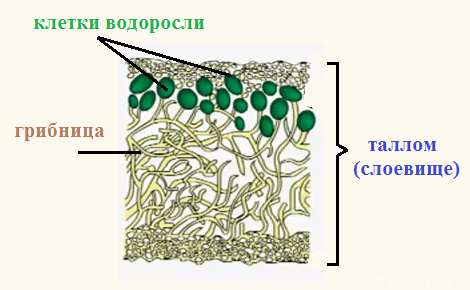
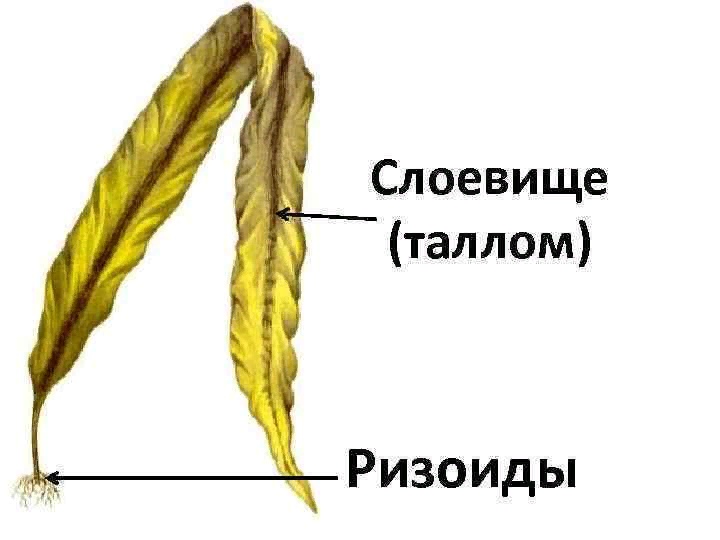
Формы слоевища у разных организмов
Слоевище водорослей называется также талломом. Он может быть нитчатый, пластинчатый, сифональный, кустистый, округлый, лентовидный, стреловидный. Форма слоевища этих организмов бывает достаточно разнообразной не только по форме, но и по цвету: тело водорослей может быть зеленым, бурым, красным, желтым или желто-зеленым, коричневым.
Лишайники также имеют различные формы своего таллома. Они могут быть:
- Накипными: таллом плотно срастается с субстратом, на котором он произрастает, и тогда тело лишайника невозможно оторвать, не повредив структуру этого субстрата.
- Листоватыми. Их отличие заключается в небольшом количестве отходящих из одной точки лопастей, которые легко отрываются от субстрата.
- Кустистыми. Они могут достигать больших размеров в длину, часто бывают хорошо и густо разветвленными.
Среди грибов также существуют талломные формы, однако у этого царства наличие групп организмов, для которых характерно слоевище, является, скорее, исключением, нежели правилом. Для них свойственно образование тела мицелием.
Талломы водорослей
Талломы водорослей весьма разнообразны, в альгологии различают несколько типов морфологической дифференциации таллома. Часто, однако, чёткую границу между различными типами провести невозможно, так как существуют переходные формы.
- Монадный, или жгутиковый тип — отдельные клетки, имеющие постоянную форму, способные к активному движению в водной среде при помощи жгутиков, одиночные или собирающиеся в колонии, ценобии (колонии с неизменным определённым числом клеток). Целые колонии и ценобии могут быть подвижными. У монадных водорослей часто встречаются характерные органеллы: центриоли, глоточный аппарат, пищеварительные вакуоли, стигма, сократительные вакуоли (у пресноводных). У многих отсутствуют вакуоли с клеточным соком и клеточная стенка, покровы монадных клеток разнообразны и отличаются от покровов клеток растений. Монадную структуру имеют также отдельные стадии водорослей (зооспоры, гаметы), обладающих многоклеточным талломом. Монадная организация таллома встречается у большинства таксонов водорослей, кроме хлорарахниофитовых, красных водорослей и цианобактерий.
- Амёбоидный, или ризоподиальный тип менее распространён, чем монадный. Амёбоидные клетки тоже могут быть одиночными или образовывать колонии, они лишены жёстких покровов, могут изменять свою форму и способны к амёбоидному движению. Встречается у хлорарахниофитовых, динофитовых и охрофитовых водорослей.
- Гемимонадный (пальмеллоидный, капсальный, трансгрессивный) тип — клетки, подобные монадным и имеющие характерные для монадного типа органеллы, но ведущие неподвижный образ жизни. Часто образуют колонии, состоящие из слизистых выделений и погружённых в эту слизь клеток. Встречается у зелёных, охрофитовых и глаукофитовых водорослей.
- Коккоидные клетки — одиночные или колониальные (ценобиальные), одетые в плотные оболочки, без жгутиков, в вегетативном состоянии неподвижные. Талломы коккоидного типа широко распространены у различных систематических групп водорослей, отсутствует этот тип у эвгленовых, гаптофитовых и криптофитовых.
- Сарциноидный тип является переходным между одноклеточными коккоидными и многоклеточными (паренхиматозными) водорослями. Клетки такого типа способны делиться в трёх взаимно перпендикулярных плоскостях и образовывать объёмные скопления. Как и для предыдущих типов, известны одиночные и колониальные представители сарциноидного типа. Насколько этот тип распространён в различных отделах водорослей, ещё окончательно не выяснено, характерные представители встречаются у зелёных водорослей, возможно также у класса жёлто-зелёных водорослей (отдел Охрофиты).
- Нитчатый, или трихальный тип — многоклеточный таллом, клетки которого делятся преимущественно в одной плоскости и образуют нити толщиной в один или несколько рядов. Нити могут быть простыми или разветвлёнными. Чаще всего клетки нити делятся поперечными перегородками, что обеспечивает постоянный рост нити в длину. Трихальный тип таллома широко представлен у охрофитовых, зелёных, красных водорослей и у цианобактерий.
- Разнонитчатый (гетеротрихальный) таллом схож с ничтатым, но различные участки его морфологически дифференцированы и приспособлены к выполнению различных функций, например, могут иметься две системы нитей — стелющиеся по субстрату и отходящие от них вертикально. Разнонитчатые талломы встречаются у охрофитовых, зелёных и красных водорослей.
- Ложнотканевый, или псевдопаренхиматозный таллом представляет собой объёмистое слоевище, возникающее в результате срастания нитей. Встречается у охрофитовых и красных водорослей (возможно, также и у зелёных).
- Тканевый, пластинчатый, или паренхиматозный тип образуется за счёт деления клеток в двух или трёх направлениях. Образуются объёмные или пластинчатые, листовидные слоевища, дифференцированные на ткани, которые выполняют различные функции. Тканевые слоевища чаще всего встречаются у охрофитов (в частности, у бурых водорослей), известны они и у представителей зелёных и красных водорослей.
- Сифональный, или сифоновый тип — это многоядерное слоевище неклеточного строения. Оно может вырастать до макроскопических размеров и иметь определённую степень дифференцировки (внешнюю расчленённость). Перегородки появляются при повреждении такого таллома и в процессе размножения. Сифональные талломы известны у зелёных и жёлто-зелёных водорослей.
- Сифонокладальный таллом образуется из первичного сифонального и представляет собой сложное слоевище нитчатой или другой формы, состоящее из многоядерных элементов. Встречается только у зелёных водорослей.
Талломы грибов
У грибов и грибоподобных организмов (псевдогрибов) обычно выделяют талломы амёбоидные и плазмодиальные, мицелиальные и дрожжеподобные.
-
Амёбоидный таллом представляет собой одноядерную клетку, не имеющую клеточной стенки (имеется лишь цитоплазматическая мембрана и перипласт — уплотнённый наружный белковый слой). Плазмодиальный таллом имеет аналогичное строение, но содержит множество клеточных ядер. По морфологическим особенностям различают талломы:
- экстраматрикальные — находящиеся на поверхности и интраматрикальные — развивающиеся внутри субстрата;
- моноцентрические — представленные одной амёбоидной одноядерной клеткой, которая может иметь короткие безъядерные корнеподобные выросты (ризомицелий) и полицентрические — с удлинённым ризомицелием, образующим утолщения, содержащие дополнительные клеточные ядра;
- холокарпические — при развитии и переходе к стадии спороношения целиком превращающиеся в один спорангий и эукарпические — формирующие несколько спорангиев.
- Мицелиальный таллом состоит из тонких ветвящихся нитей — гиф, которые могут быть разделены септами на клетки (клеточный мицелий) или не иметь септ (неклеточный мицелий). Обычно гифы имеют неограниченный верхушечный рост, характерный как для первичной гифы, так и для боковых ветвей; гифы переплетаются и образуют анастомозы — слияния боковых ветвей; поэтому в зрелом мицелии обычно невозможно различить первичную гифу и ответвления. Из мицелия формируются почти все известные грибные структуры — плодовые тела, покоящиеся видоизменения (склероции, хламидоспоры), проводящие и поддерживающие структуры (синнемы, тяжи, ризоморфы), инфекционные структуры (аппрессории, гаустории, ловчие гифы).
- Дрожжеподобный таллом (см. Дрожжи) — отдельные клетки, имеющие клеточные стенки или короткие, легко распадающиеся цепочки клеток — псевдомицелий.
Талломы водорослей
Талломы водорослей весьма разнообразны, в альгологии различают несколько типов морфологической дифференциации таллома. Часто, однако, чёткую границу между различными типами провести невозможно, так как существуют переходные формы.





- Монадный, или жгутиковый тип — отдельные клетки, имеющие постоянную форму, способные к активному движению в водной среде при помощи жгутиков, одиночные или собирающиеся в колонии, ценобии (колонии с неизменным определённым числом клеток). Целые колонии и ценобии могут быть подвижными. У монадных водорослей часто встречаются характерные органеллы: центриоли, глоточный аппарат, пищеварительные вакуоли, стигма, сократительные вакуоли (у пресноводных). У многих отсутствуют вакуоли с клеточным соком и клеточная стенка, покровы монадных клеток разнообразны и отличаются от покровов клеток растений. Монадную структуру имеют также отдельные стадии водорослей (зооспоры, гаметы), обладающих многоклеточным талломом. Монадная организация таллома встречается у большинства таксонов водорослей, кроме хлорарахниофитовых, красных водорослей и цианобактерий.
- Амёбоидный, или ризоподиальный тип менее распространён, чем монадный. Амёбоидные клетки тоже могут быть одиночными или образовывать колонии, они лишены жёстких покровов, могут изменять свою форму и способны к амёбоидному движению. Встречается у хлорарахниофитовых, динофитовых и охрофитовых водорослей.
- Гемимонадный (пальмеллоидный, капсальный, трансгрессивный) тип — клетки, подобные монадным и имеющие характерные для монадного типа органеллы, но ведущие неподвижный образ жизни. Часто образуют колонии, состоящие из слизистых выделений и погружённых в эту слизь клеток. Встречается у зелёных, охрофитовых и глаукофитовых водорослей.
- Коккоидные клетки — одиночные или колониальные (ценобиальные), одетые в плотные оболочки, без жгутиков, в вегетативном состоянии неподвижные. Талломы коккоидного типа широко распространены у различных систематических групп водорослей, отсутствует этот тип у эвгленовых, гаптофитовых и криптофитовых.
- Сарциноидный тип является переходным между одноклеточными коккоидными и многоклеточными (паренхиматозными) водорослями. Клетки такого типа способны делиться в трёх взаимно перпендикулярных плоскостях и образовывать объёмные скопления. Как и для предыдущих типов, известны одиночные и колониальные представители сарциноидного типа. Насколько этот тип распространён в различных отделах водорослей, ещё окончательно не выяснено, характерные представители встречаются у зелёных водорослей, возможно также у класса жёлто-зелёных водорослей (отдел Охрофиты).
- Нитчатый, или трихальный тип — многоклеточный таллом, клетки которого делятся преимущественно в одной плоскости и образуют нити толщиной в один или несколько рядов. Нити могут быть простыми или разветвлёнными. Чаще всего клетки нити делятся поперечными перегородками, что обеспечивает постоянный рост нити в длину. Трихальный тип таллома широко представлен у охрофитовых, зелёных, красных водорослей и у цианобактерий.
- Разнонитчатый (гетеротрихальный) таллом схож с ничтатым, но различные участки его морфологически дифференцированы и приспособлены к выполнению различных функций, например, могут иметься две системы нитей — стелющиеся по субстрату и отходящие от них вертикально. Разнонитчатые талломы встречаются у охрофитовых, зелёных и красных водорослей.
- Ложнотканевый, или псевдопаренхиматозный таллом представляет собой объёмистое слоевище, возникающее в результате срастания нитей. Встречается у охрофитовых и красных водорослей (возможно, также и у зелёных).
- Тканевый, пластинчатый, или паренхиматозный тип образуется за счёт деления клеток в двух или трёх направлениях. Образуются объёмные или пластинчатые, листовидные слоевища, дифференцированные на ткани, которые выполняют различные функции. Тканевые слоевища чаще всего встречаются у охрофитов (в частности, у бурых водорослей), известны они и у представителей зелёных и красных водорослей.
- Сифональный, или сифоновый тип — это многоядерное слоевище неклеточного строения. Оно может вырастать до макроскопических размеров и иметь определённую степень дифференцировки (внешнюю расчленённость). Перегородки появляются при повреждении такого таллома и в процессе размножения. Сифональные талломы известны у зелёных и жёлто-зелёных водорослей.
- Сифонокладальный таллом образуется из первичного сифонального и представляет собой сложное слоевище нитчатой или другой формы, состоящее из многоядерных элементов. Встречается только у зелёных водорослей.