[править] Введение
Рецептор — воспринимающий элемент; чаще всего — в биологии. Это сложное образование, состоящие из терминалей (нервных окончаний) дендритов чувствительных нейронов, глии, специализированных образований межклеточного вещества и специализированных клеток других тканей, которые в комплексе обеспечивают превращение влияния факторов внешней или внутренней среды (раздражитель) в нервный импульс — волна возбуждения, которая распространяется по нервному волокну и служит для передачи информации от периферических рецепторных (чувствительных) окончаний к нервным центрам, внутри центральной нервной системы и от неё к исполнительным аппаратам — мышцам и железам. Они состоят из «терминалей», дендрита нейронов, глий, специализированных образований межклеточного вещества и специализированных клеток иных тканей. В некоторых рецепторах (например, вкусовых и слуховых рецепторах человека) раздражитель непосредственно воспринимается специализированными клетками эпителиального происхождения или видоизмененными нервными клетками (чувствительные элементы сетчатки), которые не генерируют нервных импульсов, а действуют на иннервирующие их нервные окончания, изменяя секрецию медиатора. В других случаях единственным клеточным элементом рецепторного комплекса является само нервное окончание, часто связанное со специальными структурами межклеточного вещества (например, тельце Пачини).
В технических науках для обозначения воспринимающего элемента чаще используется термин сенсор или датчик (от англ. sensor или чувствительный), предназначенное для преобразования спроецированного на него оптического изображения в электрический сигнал и его сканирования.
Рецепторы человека[править | править код]
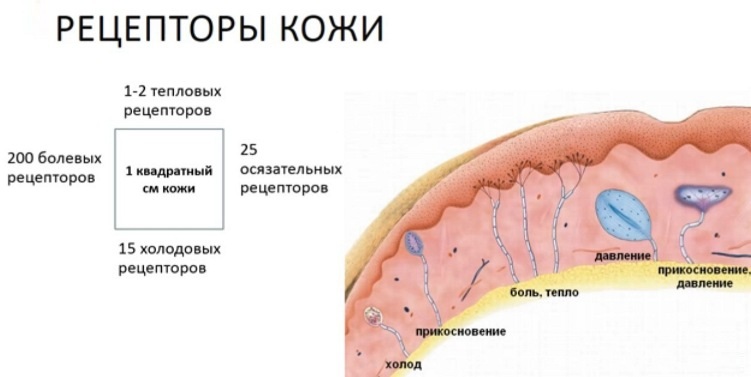
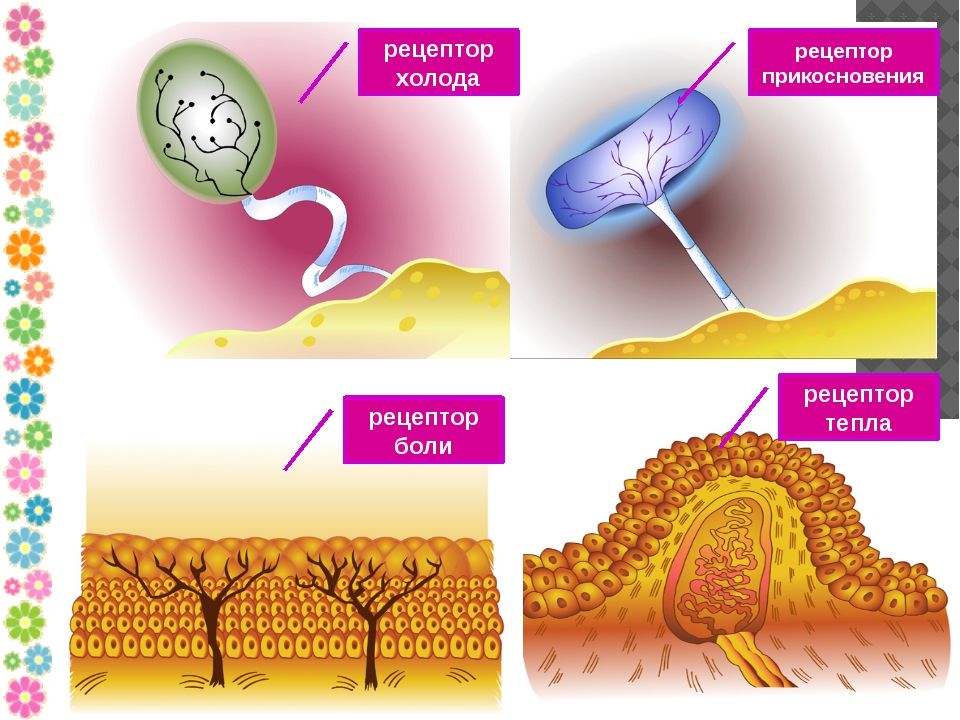
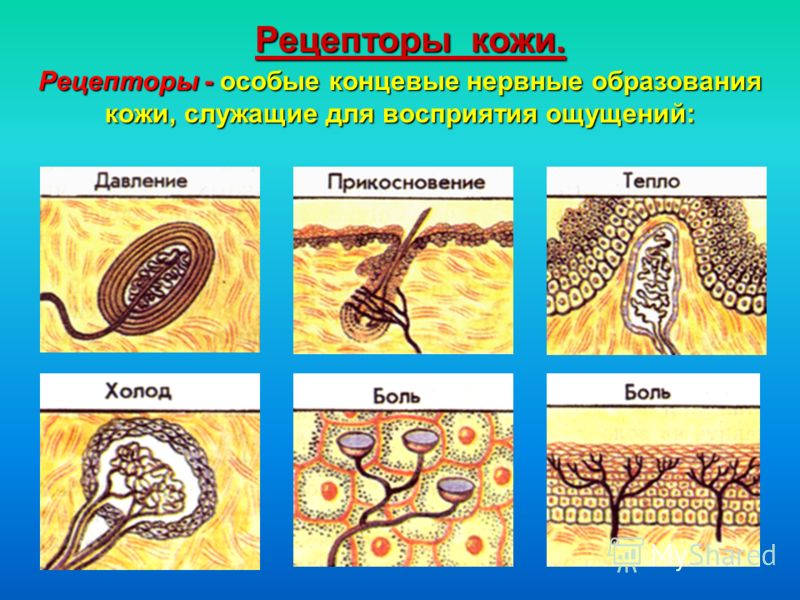
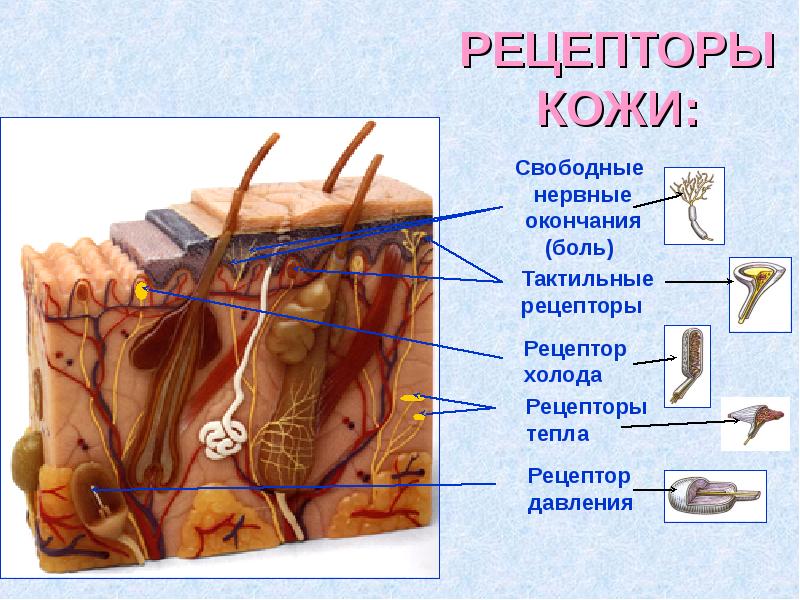
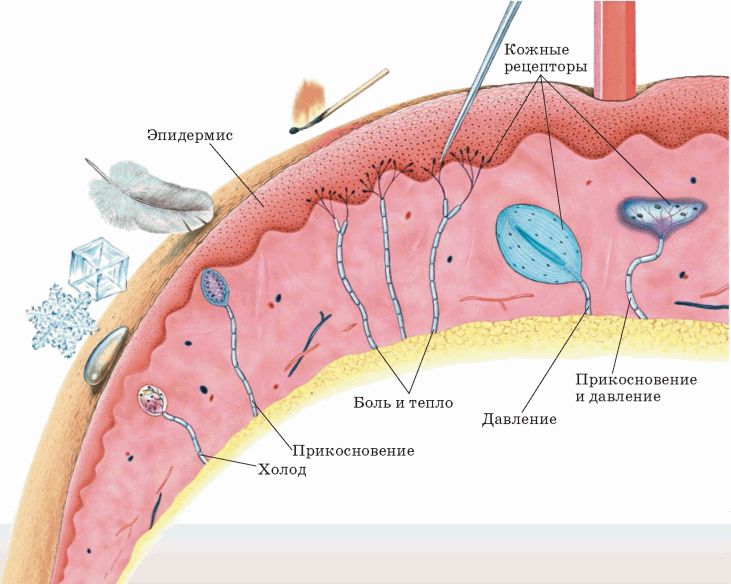
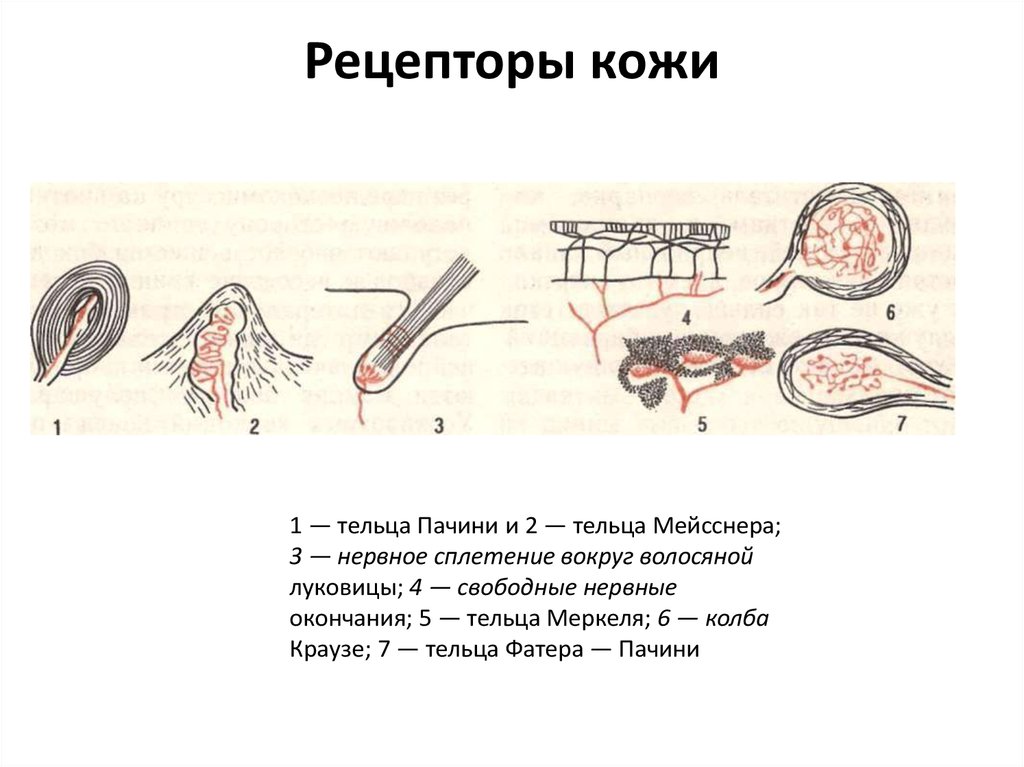
Рецепторы кожи
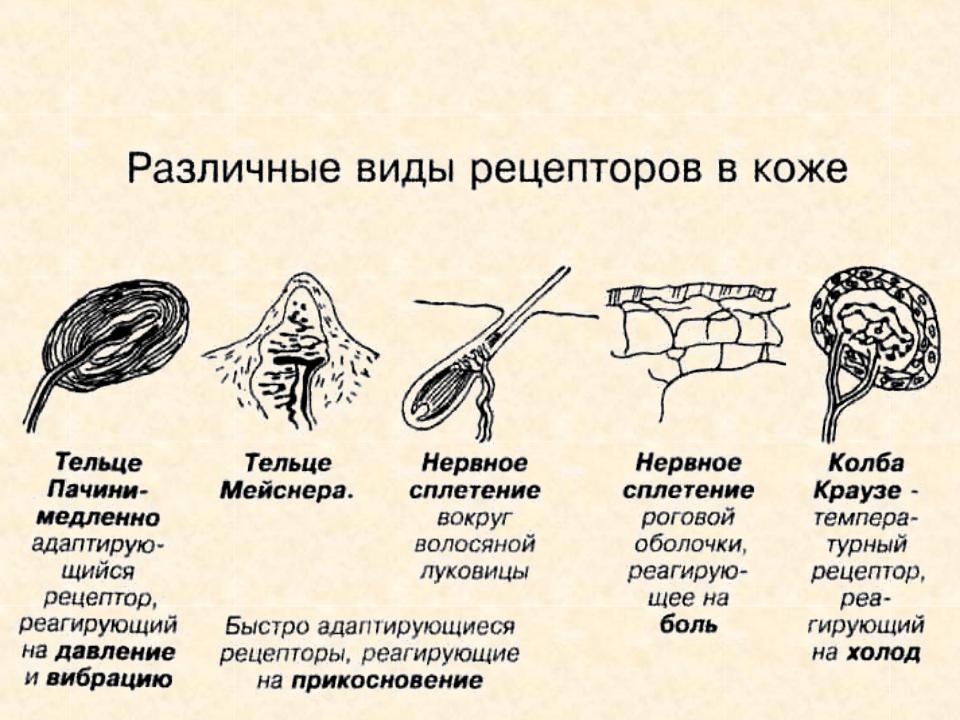
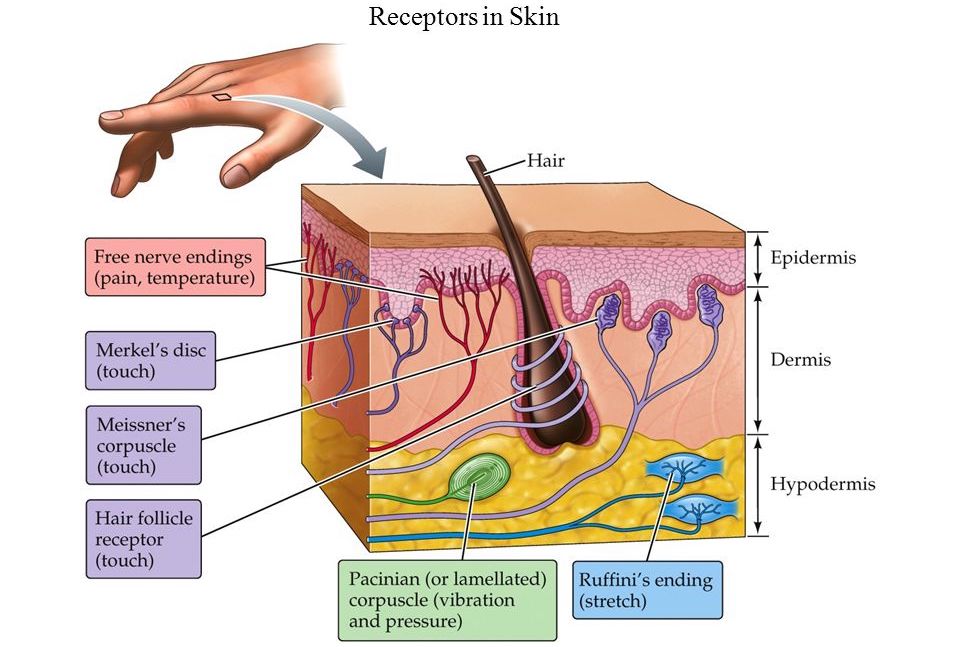
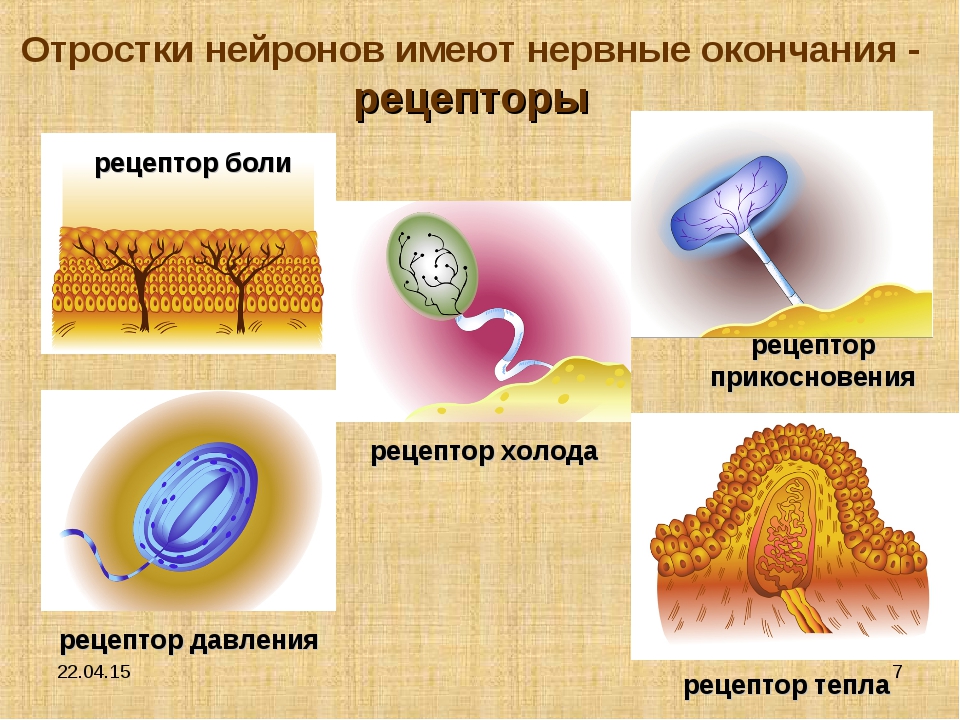
- Свободные нервные окончания — нервные окончания, состоящие только из конечных ветвлений осевого цилиндра. Располагаются в эпителии. Выступают в качестве терморецепторов, механорецепторов и ноцицепторов (то есть отвечают за восприятие изменения температуры, механических воздействий и болевые ощущения).
- Несвободные нервные окончания:
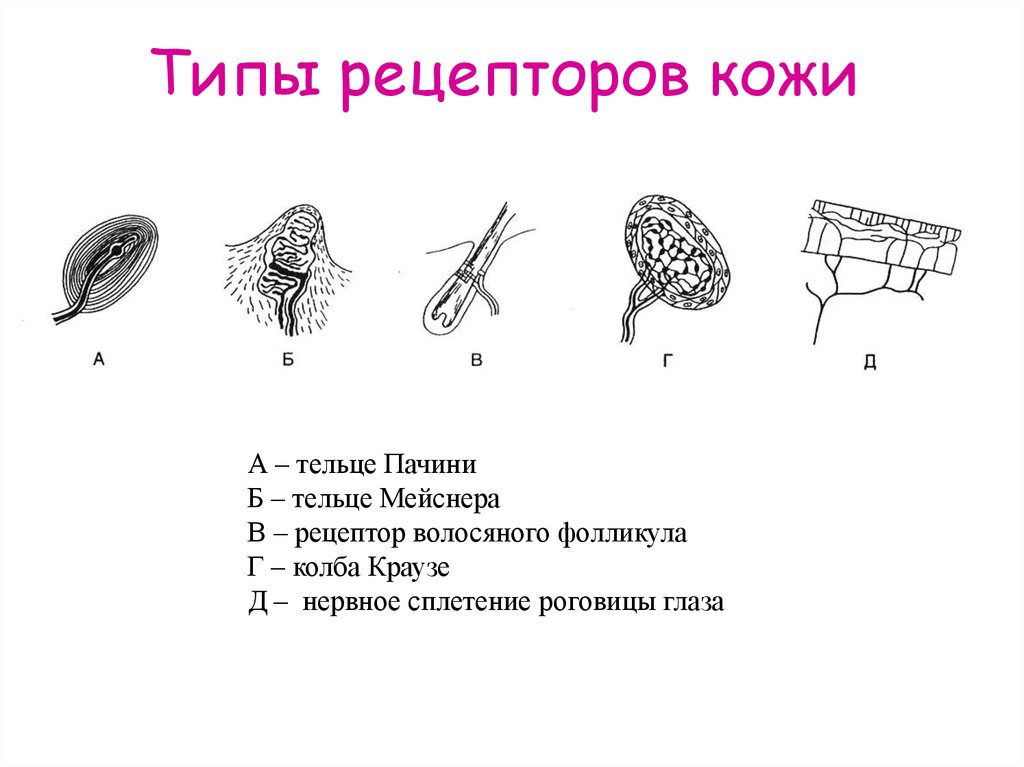
- Тельца Пачини — инкапсулированные рецепторы давления в округлой многослойной капсуле. Располагаются в подкожно-жировой клетчатке. Являются быстроадаптирующимися (реагируют только в момент начала воздействия), то есть регистрируют силу давления. Обладают большими рецептивными полями, а потому обладают грубой чувствительностью.
- Тельца Мейснера — инкапсулированные рецепторы давления, расположенные в дерме. Представляют собой слоистую структуру с нервным окончанием, проходящим между слоями. Являются быстроадаптирующимися. Обладают малыми рецептивными полями, а потому обладают тонкой чувствительностью.
- Тельца Меркеля — некапсулированные рецепторы давления. Располагаются у птиц — в дерме, у прочих позвоночных — в глубоких слоях эпидермиса. Являются медленноадаптирующимися (реагируют на всей продолжительности воздействия), то есть регистрируют продолжительность давления. Обладают малыми рецептивными полями.
- Тельца Руффини — инкапсулированные рецепторы растяжения. Являются медленноадаптирующимися, обладают большими рецептивными полями. Реагируют также на тепло.
- Колбы Краузе — инкапсулированные рецепторы, расположенные в надсосочковом слое дермы. Раньше считалось, что у Колб Краузе есть специфическая чувствительность, но их роль в качестве холодовых рецепторов не подтвердилась. .
- Рецепторы волосяных фолликулов — механорецепторы, расположенные в волосяных фолликулах и реагирующие на отклонение волоса от исходного положения.
Рецепторы мышц и сухожилий (проприоцепторы)
- Мышечные веретена — рецепторы растяжения мышц, бывают двух типов:
- с ядерной сумкой
- с ядерной цепочкой
- Сухожильный орган Гольджи — рецепторы сокращения мышц. При сокращении мышцы сухожилие растягивается и его волокна пережимают рецепторное окончание, активируя его.
Рецепторы связок
В основном представляют собой свободные нервные окончания (Типы 1, 3 и 4), меньшая группа — инкапсулированные (Тип 2). Тип 1 аналогичен окончаниям Руффини, Тип 2 — тельцам Паччини.
Рецепторы сетчатки глаза
Сетчатка содержит палочковые и колбочковые фоточувствительные клетки, в которых имеются светочувствительные пигменты. Палочки чувствительны к очень слабому свету, это длинные и тонкие клетки, сориентированные по оси прохождения света. Все палочки содержат один и тот же светочувствительный пигмент. Колбочки требуют намного более яркого освещения, это короткие конусообразные клетки, у человека колбочки делятся на три вида, каждый из которых содержит свой светочувствительный пигмент — это и есть основа цветового зрения.
Под воздействием света в рецепторах происходит выцветание — молекула зрительного пигмента поглощает фотон и превращается в другое соединение, хуже поглощающее свет на этой длине волны. Практически у всех животных (от насекомых до человека) этот пигмент состоит из белка, к которому присоединена небольшая молекула, близкая по структуре к витамину A. Эта молекула и представляет собой химически трансформируемую светом часть. Белковая часть выцветшей молекулы зрительного пигмента активирует молекулы трансдуцина, каждая из которых деактивирует сотни молекул циклического гуанозинмонофосфата, участвующих в открытии пор мембраны для ионов натрия, в результате чего поток ионов прекращается — мембрана гиперполяризуется.
Чувствительность палочек такова, что адаптировавшийся к полной темноте человек способен увидеть вспышку света такую слабую, что каждый рецептор получит не больше одного фотона. При этом палочки не способны реагировать на изменения освещённости, когда свет настолько ярок, что все натриевые каналы уже закрыты.
Лиганд (биохимия)[править | править код]
Основная статья: Лиганд (биохимия)
Рис.4. Работа лиганда.





Лаборатория клеточной биологии рецепторов РАНправить | править код
Лаборатория, оснащённая современным оборудованием, занимается изучением механизмов функционирования клеточных рецепторов. В настоящее время проводится работа по двум основным направлениям:
Первое связано с исследованием адгезионного нейронального G-белоксопряженного рецептора CIRL. Рецепторы CIRL представляют собой природные гибриды двух классов белков – сигнальных рецепторов и молекул клеточной адгезии. Считается, что эти химерные рецепторы могут быть вовлечены в межклеточные взаимодействия и передачу сигналов, опосредованных G-белками. Однако до сих пор для рецепторов данного семейства не были найдены природные агонисты.
Второе направление связано с исследованием представителя семейства инсулинового рецептора – IRR (insulin receptor-related receptor). К этому семейству принадлежат также инсулиновый рецептор (IR) и рецептор инсулино-подобного фактора роста (IGF-IR). Лигандами рецепторов IR и IGF-IR являются эндогенные пептиды, тогда как для IRR до недавнего времени не удавалось обнаружить лиганд, несмотря на значительные усилия, предпринятые в этом направлении.
Классификация рецепторов. Мономодальные и полимодальные рецепторы. Ноцицепторы ( болевые рецепторы ). Экстерорецепторы. Интерорецепторы.
Большинство рецепторов возбуждаются в ответ на действие стимулов только одной физической природы и поэтому относятся к мономодальным. Их можно возбудить и некоторыми неадекватными раздражителями, например фоторецепторы — сильным давлением на глазное яблоко, а вкусовые рецепторы — прикосновением языка к контактам гальванической батареи, но получить качественно различаемые ощущения в таких случаях невозможно. Наряду с мономодальными существуют полимодальные рецепторы, адекватными стимулами которых могут служить раздражители разной природы. К такому типу рецепторов принадлежат некоторые болевые рецепторы, или ноцицепторы (лат. nocens — вредный), которые можно возбудить механическими, термическими и химическими стимулами. Полимодальность имеется у терморецепторов, реагирующих на повышение концентрации калия во внеклеточном пространстве так же, как на повышение температуры.
В зависимости от строения рецепторов их подразделяют на первичные, или первичночувствующие, которые являются специализированными окончаниями чувствительного нейрона, и вторичные, или вторичночувствующие, представляющие собой клетки эпителиального происхождения, способные к образованию рецепторного потенциала в ответ на действие адекватного стимула. Первичночувствующие рецепторы могут сами генерировать потенциалы действия в ответ на раздражение адекватным стимулом, если величина их рецепторного потенциала достигнет пороговой величины. К ним относятся обонятельные рецепторы, большинство механорецепторов кожи, терморецепторы, болевые рецепторы или ноцицепторы, проприоцепторы и большинство интерорецепторов внутренних органов.
Вторичночувствующие рецепторы отвечают на действие раздражителя лишь возникновением рецепторного потенциала, от величины которого зависит количество выделяемого этими клетками медиатора. С его помощью вторичные рецепторы действуют на нервные окончания чувствительных нейронов, генерирующих потенциалы действия в зависимости от количества медиатора, выделившегося из вторичночувствующих рецепторов. Вторичные рецепторы представлены вкусовыми, слуховыми и вестибулярными рецепторами, а также хемочувствительными клетками синокаротидного клубочка. Фоторецепторы сетчатки, имеющие общее происхождение с нервными клетками, чаще относят к первичным рецепторам, но отсутствие у них способности генерировать потенциалы действия указывает на их сходство с вторичными рецепторами.




В зависимости от источника адекватных стимулов рецепторы подразделяют на наружные и внутренние, или экстерорецепторы и интерорецепторы; первые стимулируются при действии раздражителей внешней среды (электромагнитные и звуковые волны, давление, действие пахучих молекул), а вторые — внутренней (к этому типу рецепторов относят не только висцерорецепторы внутренних органов, но также проприоцепторы и вестибулярные рецепторы). В зависимости от того, действует стимул на расстоянии или непосредственно на рецепторы, их подразделяют еще на дистантные и контактные.
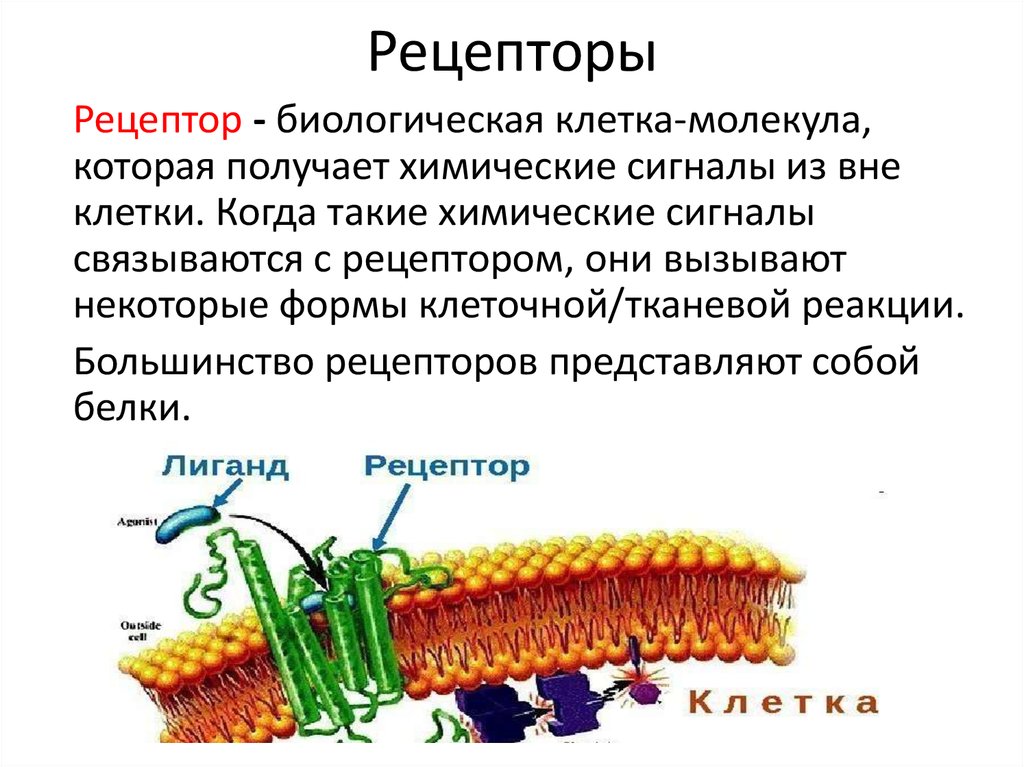
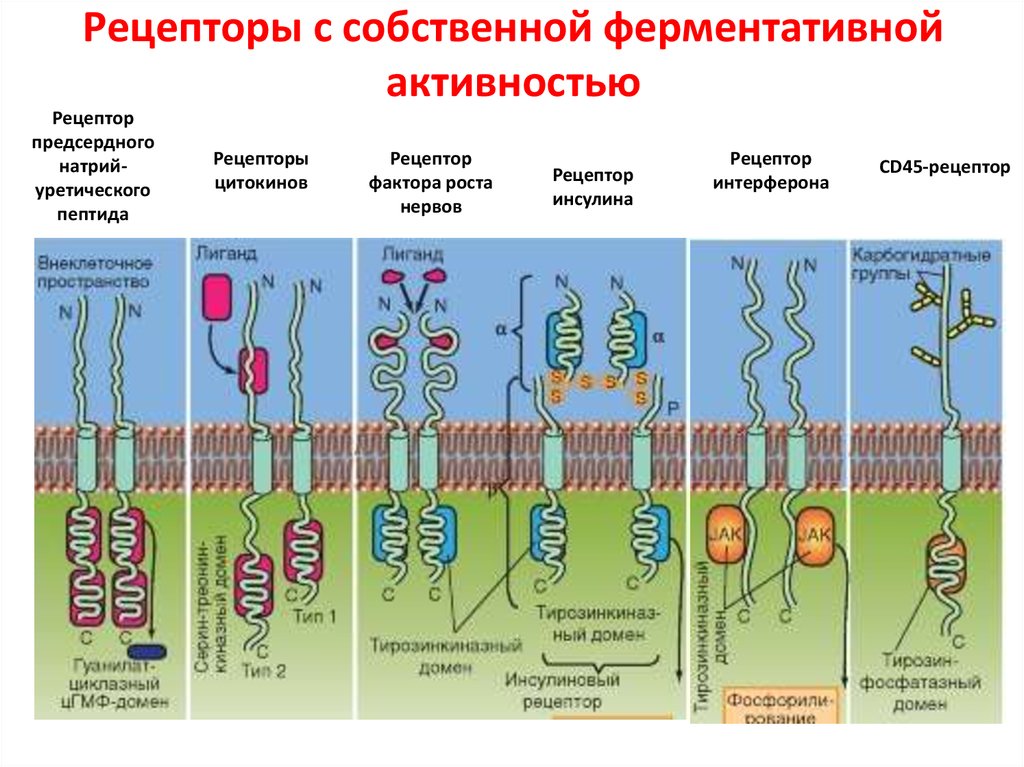
Клеточные рецепторы
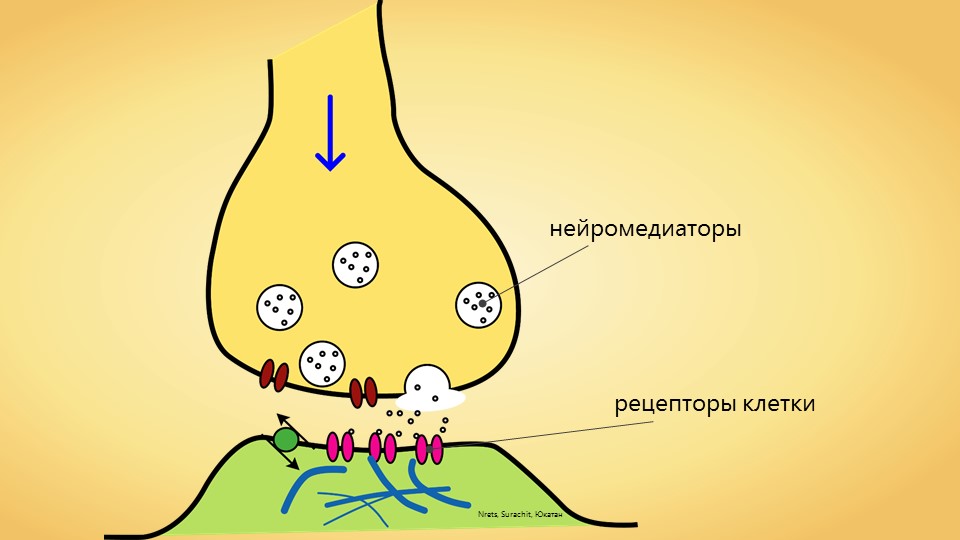
В многоклеточном организме передача информации между клетками, происходящая с участием гормонов, нейротрансмиттеров (медиаторов), нейропептидов и других биологически активных веществ, включает этап взаимодействия молекул этих веществ (их называют также лигандами) с соответствующими надмолекулярными структурами, или клеточными рецепторами. Они могут располагаться как внутри клетки (напр., клеточные Р. к стероидным гормонам, легко проникающим внутрь клетки благодаря их растворимости в липидах клеточной мембраны), так и на поверхности клеточной мембраны (клеточные Р. к белкам, пептидам, нейротрансмиттерам). Как внутриклеточные, так и мембранные клеточные Р. содержат центр связывания, обеспечивающий специфическое связывание лиганда с клеточными Р. После связывания, напр., молекулы стероидного гормона с цитоплазматическим Р. и образования комплекса гормон — клеточный Р. этот комплекс проникает внутрь клеточного ядра, где связывается с соответствующим акцептором, вслед за чем молекула гормона отделяется от комплекса и выходит в цитоплазму, при этом одновременно активируется генетический аппарат клетки (см.). Конечным итогом этой активации является резкое усиление синтеза ряда специфических и неспецифических белков клетки, что представляет собой ответ клетки-мишени на действие гормона.
Процессы, происходящие при связывании молекулы лиганда с клеточным Р., локализованным на клеточной мембране (см. Мембраны биологические), состоят из ряда этапов, протекающих с большой скоростью. Происходящее при этом изменение свойств фосфолипидного матрикса, окружающего клеточный Р., обеспечивает передачу сигнала от центра связывания лиганда (через ряд промежуточных звеньев) на аденилат-циклазный центр и его активацию. Циклическая АМФ (см. Аденозинфосфорные кислоты) является своеобразным вторым внутриклеточным переносчиком информации, определяющим ответ клетки на действие лиганда. Т. о., происходит активация соответствующих протеинкиназ, изменение проницаемости клеточной мембраны для ряда ионов, усиление экспрессии генетической информации. Важным открытием явилось обнаружение в ц. н. с. клеточных Р. к ряду нейрональных пептидов, напр, к группам пептидов, названных эндорфинами и энкефалинами, а также клеточных Р. ко многим психотропным лекарственным препаратам (имипрамину, галоперидолу, диазепаму и др.). Взаимодействие указанных выше лигандов с клеточными Р. изменяет способность нервных клеток отвечать на действие нейро-трансмиттеров, т. е. оказывает на их активность модулирующее действие. Напр., связывание препаратов бензодиазепинового ряда с бензодиазе-пиновыми клеточными Р. усиливает ответ ГАМК-ергических нейронов на действие гамма-аминомасляной к-ты (ГАМК), влияя в то же время на связывание ГАМК соответствующими клеточными Р. В связи с обнаружением в ц. н. с. эндогенных лигандов, конкурирующих с морфином за места связывания и обладающих морфиноподобным действием, проводится поиск эндогенных соединений типа эндогенного диазепама, эндогенного галоперидола и др., что может иметь в случае их обнаружения большое значение для клин, практики.
Нарушения механизмов рецепции играют важную роль в развитии ряда заболеваний человека, напр, нек-рых видов сахарного диабета, гиперхоли-стеринемии и др. Наряду с рассмотренными выше видами Р. на поверхности В- и Т-лимфоцитов выявлены мембранные клеточные Р., играющие важную роль в работе иммунной системы, а также клеточные Р. к ряду вирусов.
Библиография: Глебов Р. И. и Крыжановский Г. Н. Функциональная биохимия синапсов, М., 1978; Гранит Р. Электрофизиологическое исследование рецепции, пер. с англ., М., 1957; Розен В. Б. и Смирнов А. Н. Рецепторы и стероидные гормоны, М., 1981, библиогр.; Тамар Г. Основы сенсорной физиологии, пер. с англ., М., 1976; Физиология сенсорных систем, под ред. А. С. Батуева, с. 34, Л., 1976; Cell membrane receptors for drugs and hormones, a multidisciplinary approach, ed. by R. W. Straub a. L. Bolis, N. Y., 1978; Cell! membrane receptors for viruses, antigens, and antibodies, polypeptide hormones, and small molecules, ed. by R. F. Beers a. E. G. Bassett, N. Y., 1976; The receptors, a comprehensive treatise, ed. by R. D. O’Brien, v. 1, N. Y.— L., 1979.
Формы и действия рецепторов[править | править код]
Как потенциал действия перемещается вдоль аксона, происходит изменение полярности через мембраны. Отель na+ и K+ охраняющая ионные каналы, открываются и закрываются как мембраны достигает порогового потенциала, в ответ на сигнал от другого нейрона. В начале потенциала действия на Nа+ каналов и na+ движется внутрь аксона, вызывая деполяризацию. Реполяризация происходит, когда на K+ каналы и K+ выходит из аксона. Это создает изменение полярности между вне клетки и внутри. Импульс проходит вниз по аксону в одном направлении, в терминале аксона, где это сигналы касаются других нейронов.

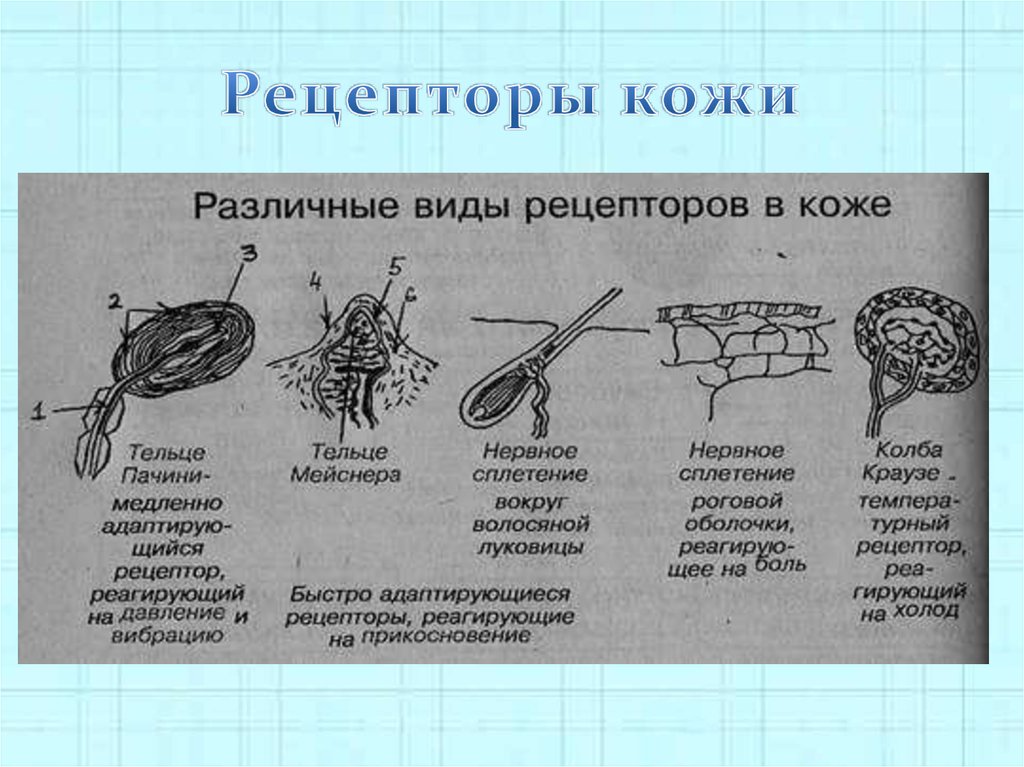

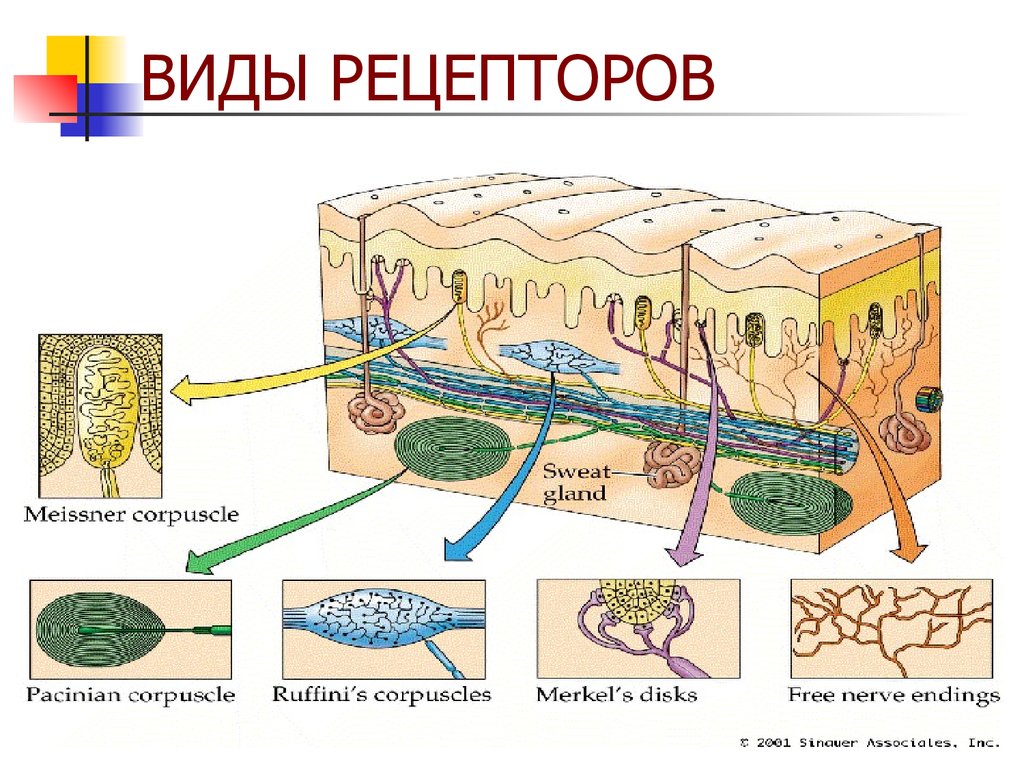




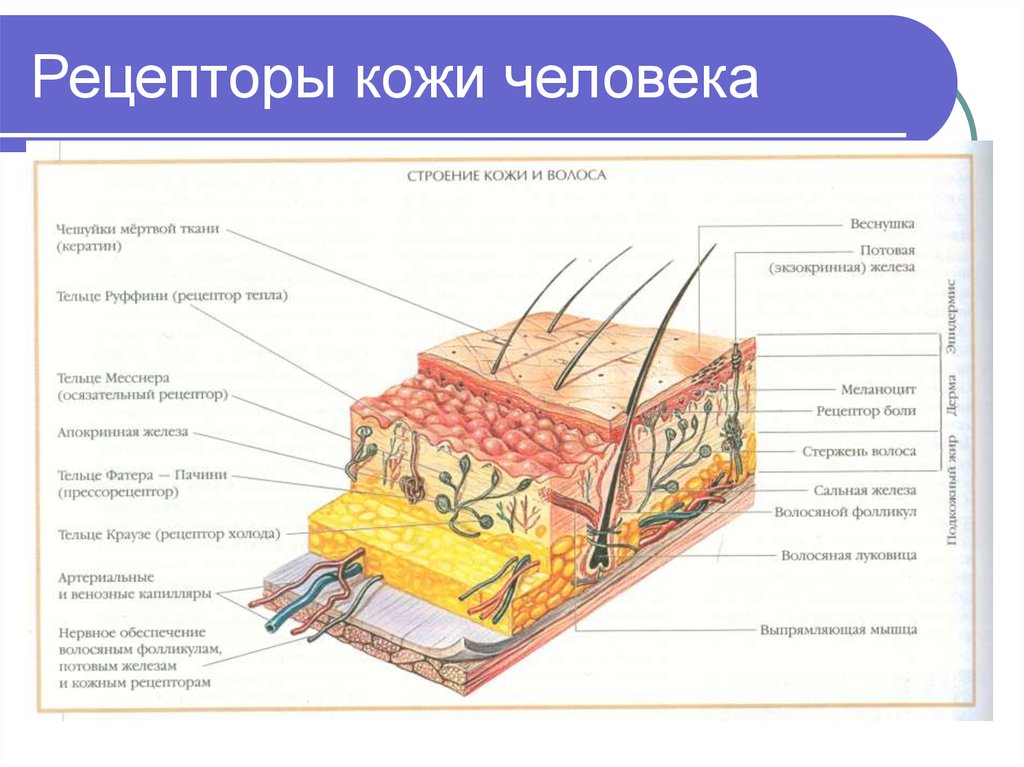

Рис. 1. Схема распределения зарядов по разные стороны мембраны возбудимой клетки в спокойном состоянии (A) и при возникновении потенциала действия (B) (см. объяснения в тексте Потенциал действия).
Формы и действия рецепторов изучены рентгеновской кристаллографией, двойной polarisation интерферометрией, компьютерным моделированием, и исследованияи функции структуры, которые продвинули понимание действия препарата на обязательных участках рецепторов. Коррелят отношений деятельности структуры вызвал конформационные изменения с биомолекулярной деятельностью, и изучен, используя динамические методы, типа круглого дихроизма и двойной polarisation интерферометрии.
Трансмембранное место — receptor: E = extracellular ; место I = intracellular; мембрана P = plasma. В зависит от их функций и лигандов, которые могут идентифицировать несколько типов рецепторов (см. рис.1t):
- Некоторые белки рецептора — периферийные мембранные белки.
- Много гормонов и рецепторов медиатора — трансмембранные белки: трансмембранные рецепторы вложены в двойной слой фосфолипида мембран ячейки, которые позволяют активацию троп трансдукции сигнала в ответ на активацию обязательной молекулой, или лигандом.
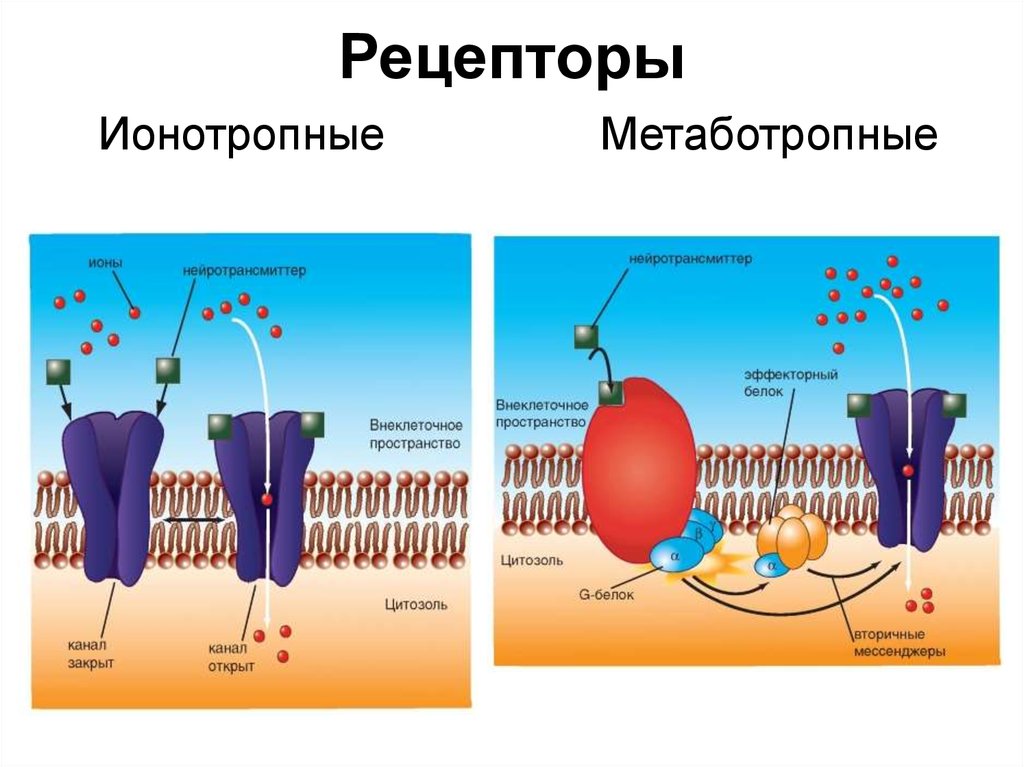
- Рецепторы Metabotropic соединены с белками Г и затрагивают ячейку косвенно через ферменты, которые управляют каналами иона.
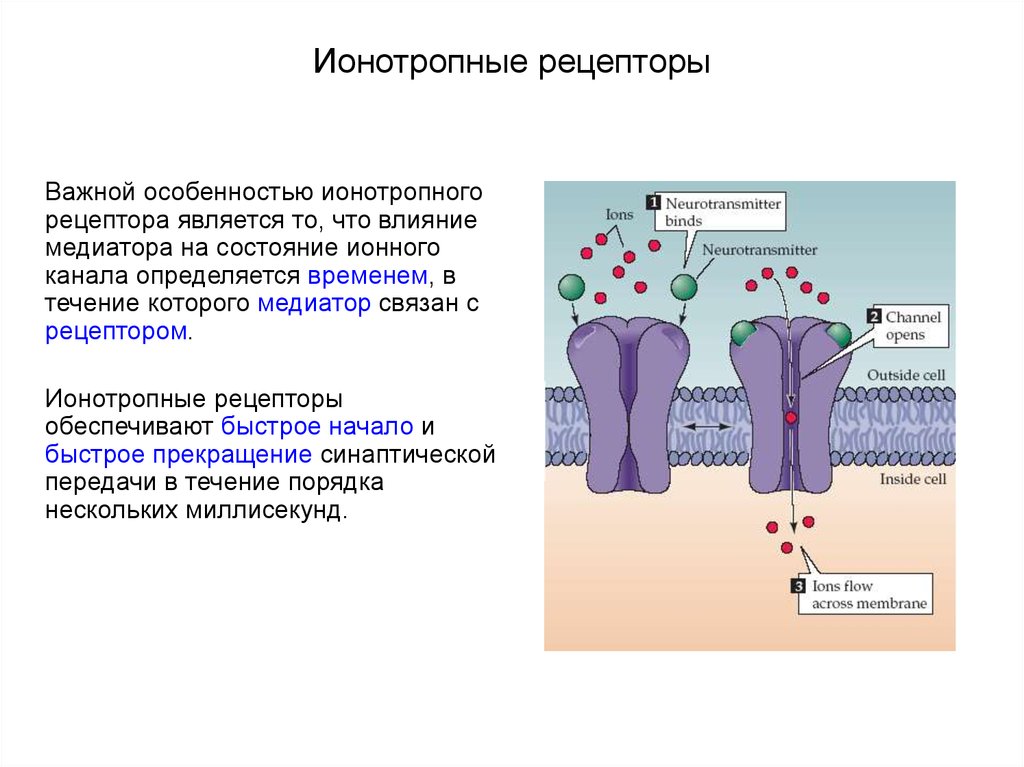
- Рецепторы Ionotropic (также известные как каналы иона лиганда-gated) содержат центральную пору, которая открывается в ответ на закрепление лиганда.
- Другой главный класс рецепторов — внутриклеточные белки, типа тех, которые для стероида и intracrine гормональных рецепторов пептида. Эти рецепторы часто могут входить в ядро ячейки и модулировать генное выражение в ответ на активацию лигандом.
Мембранные рецепторы изолированы от мембран ячейки в соответствии со сложными процедурами извлечения, используя растворители, моющие средства, и/или очистка близости.
Стимулами для разных рецепторов могут служить свет, механическая деформация — изменение размеров, химические вещества, изменения температуры, а также изменения электрического и магнитного поля. В рецепторных клетках (будь то непостредственно нервные окончания или специализированные клетки) соответствующий сигнал изменяет конформацию чувствительных молекул-клеточных рецепторов, что приводит к изменению активности мембранных ионных рецепторов и к изменению мембранного потенциала клетки. Если воспринимающей клеткой является непосредственно нервное окончание (так называемые первичные рецепторы), то происходит деполяризация мембраны с последующей генерацией нервного импульса. Специализированные рецепторные клетки вторичных рецепторов могут как де-, так и гиперполяризоваться. В последнем случае изменение мембранного потенциала ведет к уменьшению секреции тормозного медиатора, действующего на нервное окончание и, в конечном счете, все равно к генерации нервного импульса. Напаример, вызвать генерацию нервного импульса может фотон, попавший на мембрану клетки находящейся в фоточувствительном слое сетчатки.
В качестве клеточных рецепторных молекул могут выступать либо механо-, термо- и хемочувствительные ионные каналы, либо специализированные G-белки (как в клетках сетчатки). В первом случае открытие каналов непосредственно изменяет мембранный потенциал (механочувствительные каналы в тельцах Пачини), во втором случае запускается каскад внутриклеточных реакций трансдукции сигнала, что ведет в конечном счете к открытию каналов и изменению потенциала на мембране.





Виды рецепторов
Существуют несколько классификаций рецепторов:
- По положению в организме
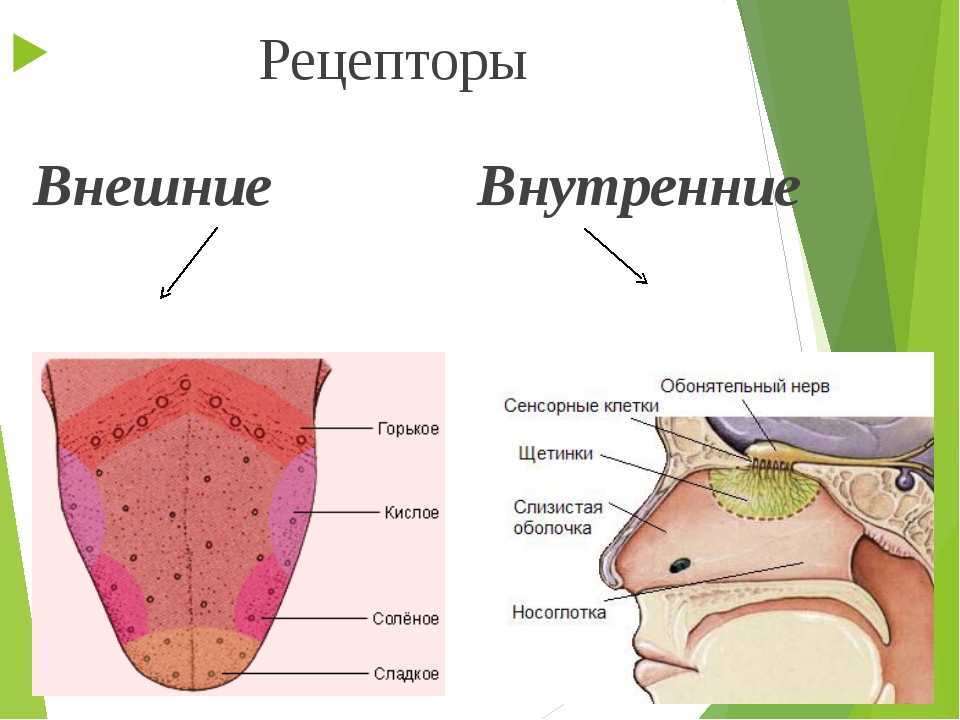
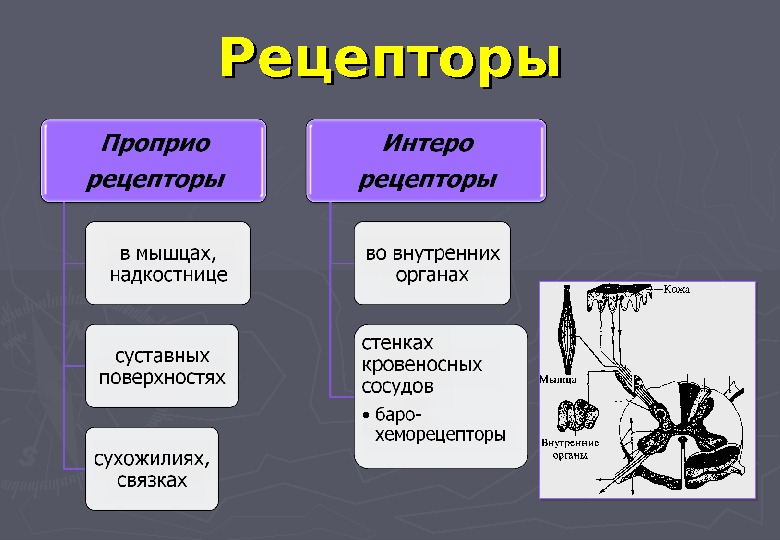
- Экстерорецепторы (экстероцепторы) — расположены на поверхности или вблизи поверхности тела и воспринимают внешние стимулы (сигналы из окружающей среды)
- Интерорецепторы (интероцепторы) — расположены во внутренних органах и воспринимают внутренние стимулы (например, информацию о состоянии внутренней среды организма)
- По способности воспринимать разные стимулы
- Мономодальные — реагирующие только на один тип раздражителей (например, фоторецепторы — на свет)
- Полимодальные — реагирующие на несколько типов раздражителей (например, многие болевые рецепторы, а также некоторые рецепторы беспозвоночных, реагирующие одновременно на механические и химические стимулы)
- По адекватному раздражителю:
- Хеморецепторы — воспринимают воздействие растворённых или летучих химических веществ
- Осморецепторы — воспринимают изменения осмотической концентрации жидкости (как правило, внутренней среды)
- Механорецепторы — воспринимают механические стимулы (прикосновение, давление, растяжение, колебания воды или воздуха и т. п.)
- Фоторецепторы — воспринимают видимый и ультрафиолетовый свет
- Терморецепторы — воспринимают понижение (холодовые) или повышение (тепловые) температуры (тепловые стимулы)
- Болевые рецепторы, стимуляция которых приводит к возникновению болевых ощущений. Такого физического стимула, как боль, не существует, поэтому выделение их в отдельную группу по природе раздражителя в некоторой степени условно. В действительности, они представляют собой высокопороговые сенсоры различных (химических, термических или механических) повреждающих факторов. Однако уникальная особенность ноцицепторов, которая не позволяет отнести их, например, к «высокопороговым терморецепторам», состоит в том, что многие из них полимодальны: одно и то же нервное окончание способно возбуждаться в ответ на несколько различных повреждающих стимулов.
- Электрорецепторы — воспринимают изменения электрического поля
- Магнитные рецепторы — воспринимают изменения магнитного поля
У человека имеются первые шесть типов рецепторов. На хеморецепции основаны вкус и обоняние, на механорецепции — осязание, слух и равновесие, а также ощущения положения тела в пространстве, на фоторецепции — зрение. Терморецепторы есть в коже и некоторых внутренних органах. Большая часть интерорецепторов запускает непроизвольные и в большинстве случаев неосознаваемые, вегетативные рефлексы. Так, осморецепторы включены в регуляцию деятельности почек, хеморецепторы, воспринимающие pH, концентрации углекислого газа и кислорода в крови, включены в регуляцию дыхания и т. д.
Иногда предлагается выделять группу электромагнитных рецепторов, в которую включают фото-, электро- и магниторецепторы. Магниторецепторы точно не идентифицированы ни у одной группы животных, хотя предположительно ими служат некоторые клетки сетчатки птиц, а возможно, и ряд других клеток.
В таблице приведены данные о некоторых типах рецепторов
| Природа раздражителя | Тип рецептора | Место расположения и комментарии |
|---|---|---|
| • электрическое поле | • ампула Лоренцини и другие типы | • Имеются у рыб, круглоротых, амфибий, а также у утконоса и ехидны |
| • химическое соединение | • хеморецептор | |
| • влажность | • гигрорецептор | • Относятся к осморецепторам или механорецепторам. Располагаются на антеннах и ротовых органах многих насекомых |
| • механическое воздействие | • механорецептор | • У человека имеются в коже (экстероцепторы) и внутренних органах (барорецепторы, проприоцепторы) |
| • давление | • барорецептор | • Относятся к механорецепторам |
| • положение тела | • проприоцептор | • Относятся к механорецепторам. У человека это нервно-мышечные веретена, сухожильные органы Гольджи и др. |
| • осмотическое давление | • осморецептор | • В основном интерорецепторы; у человека имеются в гипоталамусе, а также, вероятно, в почках, стенках желудочно-кишечного тракта, возможно, в печени. Существуют данные о широком распространении осморецепторов во всех тканях организма |
| • свет | • фоторецептор | |
| • температура | • терморецептор | • Реагируют на изменение температуры. У человека имеются в коже и в гипоталамусе |
| • повреждение тканей | • ноцицептор | • В большинстве тканей с разной частотой. Болевые рецепторы — свободные нервные окончания немиелинизированных волокон типа C или слабо миелинизированных волокон типа Aδ. |
| • магнитное поле | • магнитные рецепторы | • Точное расположение и строение неизвестны, наличие у многих групп животных доказано поведенческими экспериментами |
Виды некоторых рецепторов и их данные[править | править код]
Cм. в таблице:
| Типы рецепторов и их данные | ||
|---|---|---|
| Вид раздражителя | Тип рецептора | Комментарии |
| Электрическое поле | ||
| Хеморецептор | ||
| Относятся к осморецепторам или механорецепторам. Располагаются на антеннах и ротовых органах многих насекомых | ||
| Механорецептор | У человека имеются в коже (экстероцепторы кожи) и внутренних органах (барорецепторы, проприоцепторы) | |
| Давление. | Относятся к механорецепторам | |
| (или механорецепторы вестибулярного аппарата), реагируют на ускорения и вибрации при наклоне тела или головы. | ||
| В основном интерорецепторы; у человека имеются в гипоталамусе, а также, вероятно, в почках, стенках желудочно-кишечного тракта, возможно, в печени. Существуют данные о широком распространении осморецепторов во всех тканях организма | ||
| Свет, Цвет | Фоторецептор,Экстерорецептор | Участвуют в восприятии света и цвета |
| Температура | Реагируют на изменение температуры. У человека они имеются в коже и в гипоталамусе. | |
| Болевые рецепторы — свободные нервные окончания немиелинизированных волокон типа C или слабо миелинизированных волокон типа Aδ. | ||
| Магнитное поле | Точное расположение и строение неизвестны, наличие у многих групп животных доказано поведенческими экспериментами. |
Виды рецепторов[править | править код]
Существуют несколько классификаций рецепторов:
- По положению
- Экстерорецепторы — расположены на поверхности или вблизи поверхности тела и воспринимают внешние стимулы (сигналы из окружающей среды)
- Интерорецепторы — расположены во внутренних органах и воспринимают внутренние стимулы (например, информацию о состоянии внутренней среды организма)
- По способности воспринимать разные стимулы
- Мономодальные — реагирующие только на один тип раздражителей (например, фоторецепторы — на свет)
- Полимодальные — реагирующие на несколько типов раздражителей (например. многие болевые рецепторы, а также некоторые рецепторы беспозвоночных, реагирующие одновременно на механические и химические стимулы).
- По сенсорной модальности или адекватному раздражителю
- Хеморецепторы — воспринимают воздействие растворенных или летучих химических веществ, изменение уровня сахара в крови, рецепторы вкуса, обоняния;
-
Фоторецепторы
Экстерорецепторы сетчатки глаза;
— биологическая структура и элемент зрительной системы живых организмов, воспринимающие видимые лучи спектра прямого и/или отражённого излучения ультрафиолетового, инфракрасного и/или иного в зависимости от характеристик органа зрительной системы, в том числе и человека);
Рецепторы связокправить | править код
В основном представляют собой свободные нервные окончания (Типы 1, 3 и 4), меньшая группа — инкапсулированные (Тип 2). Тип 1 аналогичен окончаниям Руффини, Тип 2 — тельцам Паччини.