Искусственное получение мутаций
В природе постоянно идет спонтанный мутагенез, но спонтанные мутации — достаточно редкое явление, например, у дрозофилы мутация белых глаз образуется с частотой 1:100 000 гамет.
Факторы, воздействие которых на организм приводит к появлению мутаций, называются мутагенами. Обычно мутагены подразделяют на три группы. Для искусственного получения мутаций используются физические и химические мутагены.
| Название группымутагенов | Примеры |
|---|---|
| Физические | Рентгеновские лучи, гамма лучи, ультрафиолетовое излучение, высокие и низкие температуры и др. |
| Химические | Соли тяжелых металлов, алкалоиды, чужеродные ДНК и РНК, аналоги азотистых оснований нуклеиновых кислот, мн. алкилирующие соединения и др. |
| Биологические | Вирусы, бактерии |
Индуцированный мутагенез имеет большое значение, поскольку дает возможность создания ценного исходного материала для селекции, а также раскрывает пути создания средств защиты человека от действия мутагенных факторов.
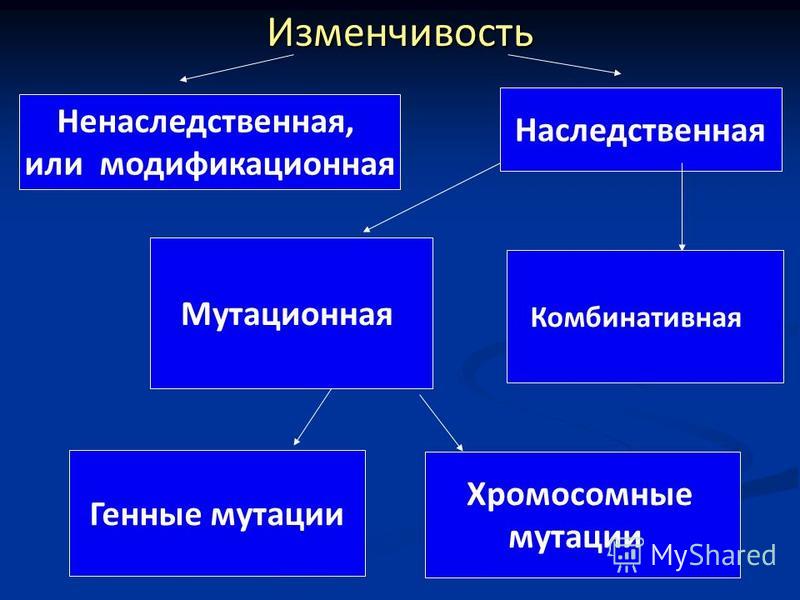
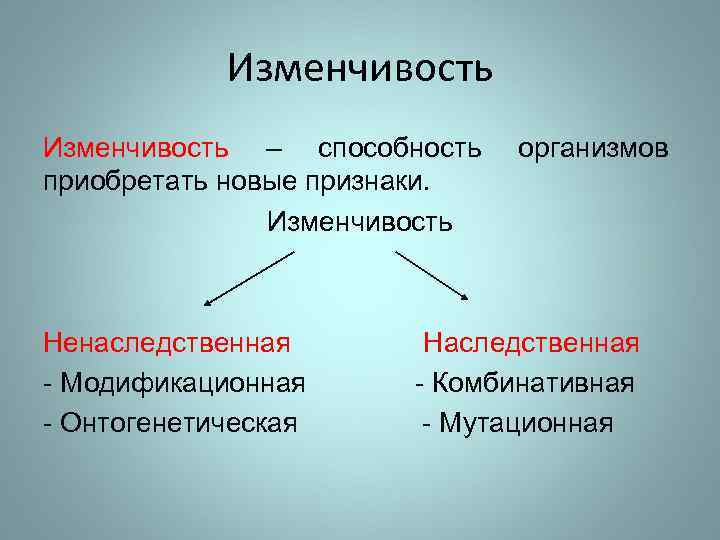
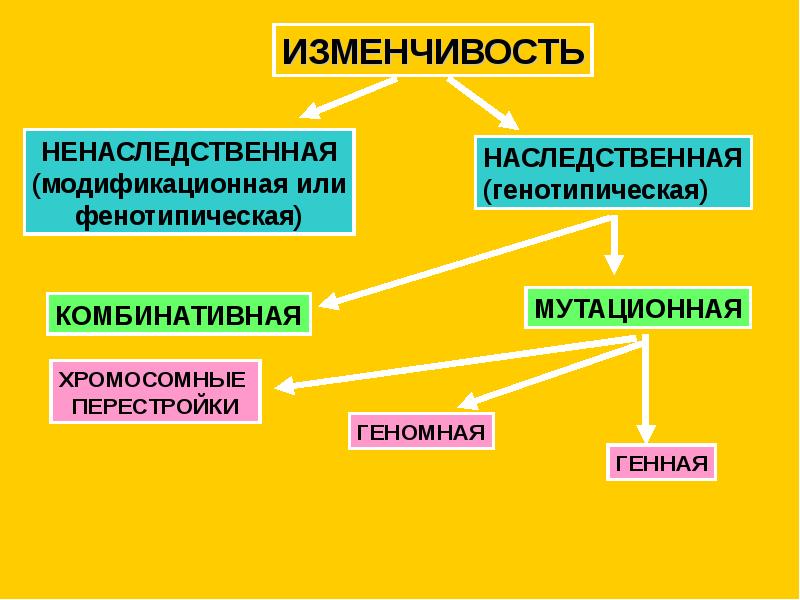
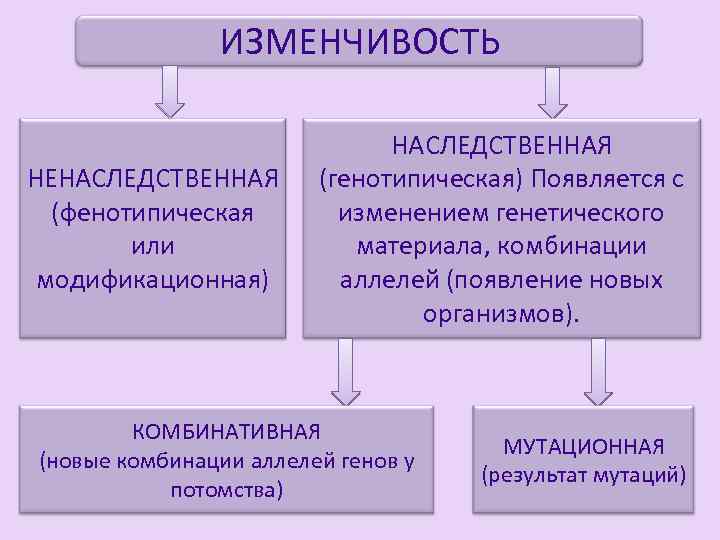
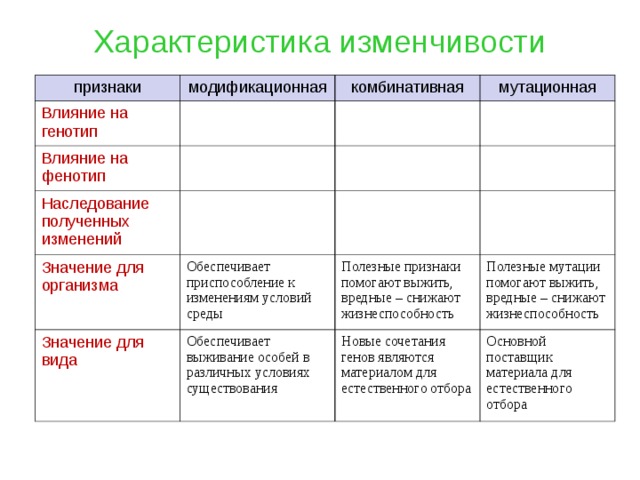
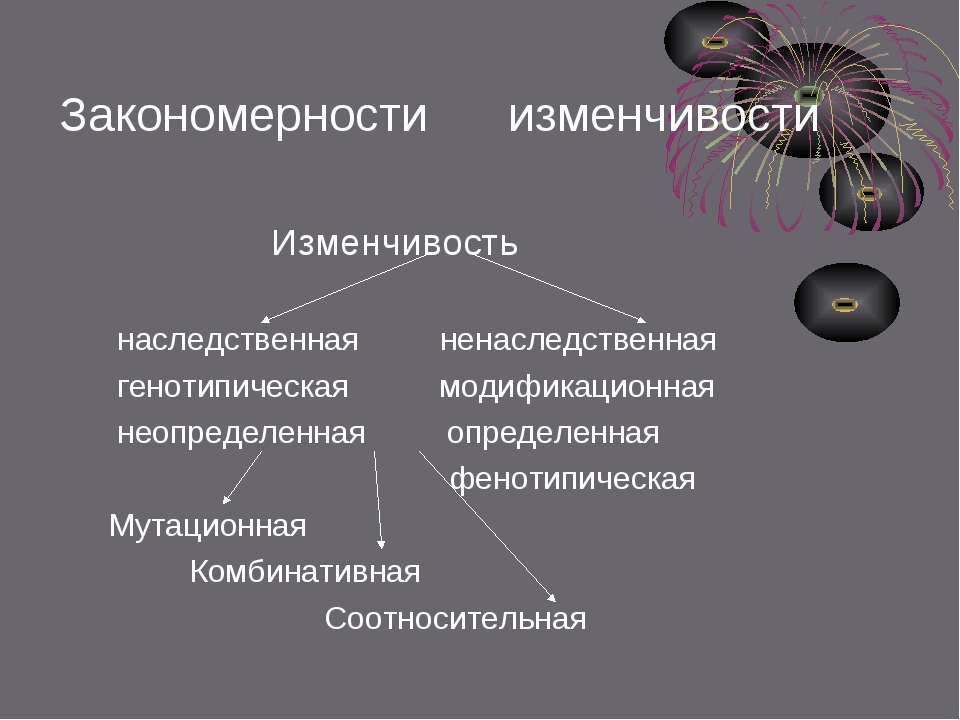
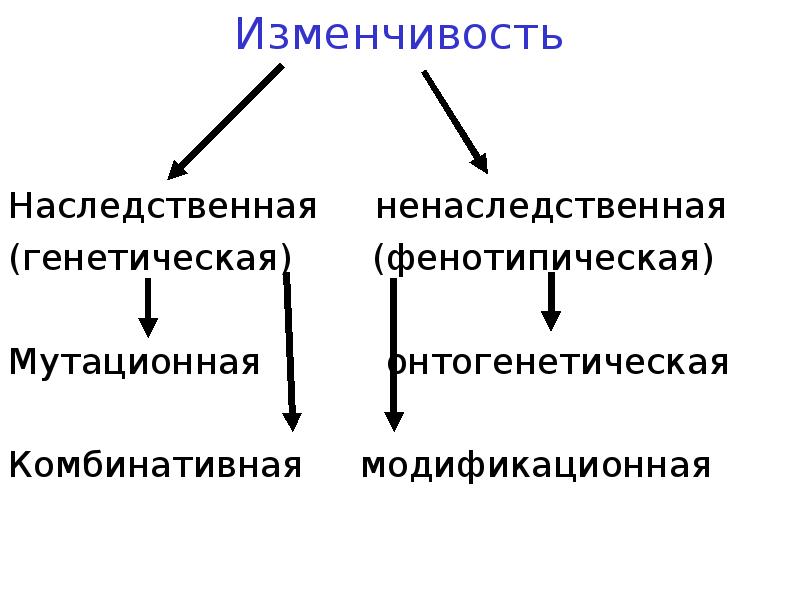
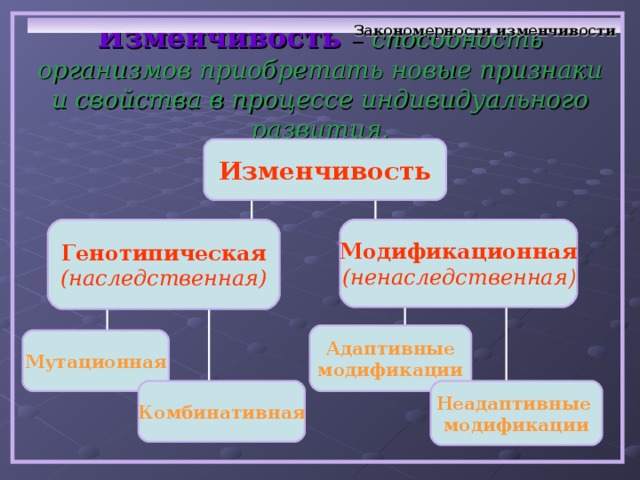
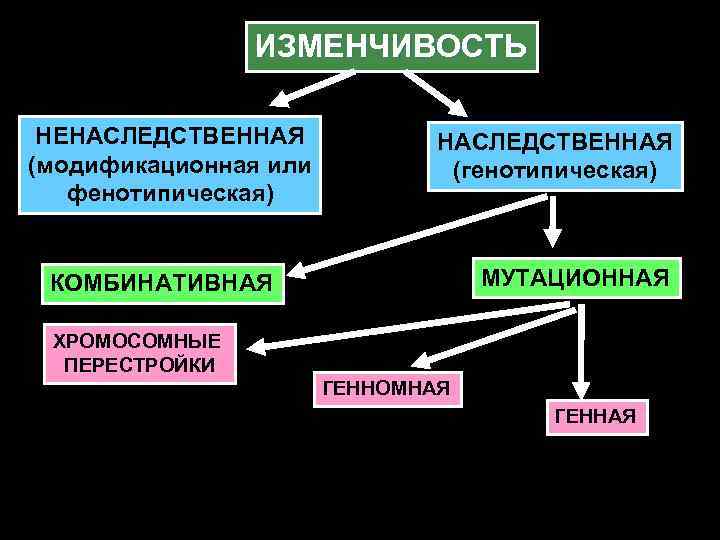
Виды мутаций и их характеристика
Мутации бывают доминантные, проявляющиеся в первом поколении и рецессивные, полезные и вредные.

Типы мутаций различают по способу возникновения:
- спонтанные или случайные, возникающие при нормальных условиях жизни и зависящие от внешних и внутренних факторов,
- индуцированные, полученные с помощью мутагенов различной природы.
По характеру проявления:
- доминантные, проявляющиеся в первом поколении,
- рецессивные, часто понижающие жизнеспособность.
По месту возникновения:
- генеративные — представляют собой мутации, возникающие в половых клетках или спорах и проявляющиеся через поколение,
- соматические, наследуемые при вегетативном размножении.
По уровню возникновения:
- генные. Их причины появления изменение геномной последовательности нуклеотидов в ДНК,
- хромосомные перестройки – изменения структуры хромосом в результате разрыва хромосомы,
- геномные – изменение числа хромосом. Геном это комплекс генов организма определенного вида.
Мутационная теория
Мутационная изменчивость была изучена в конце 19 – начале 20 веков Гуго де Фризом. Он наблюдал растение ослинник и заметил, что у него сравнительно часто возникают новые свойства. Де Фриз ввёл для обозначения этих изменений термин «мутация».
Сначала не было известно, какие структуры клетки перестраиваются при мутационной изменчивости. Но позднее было обнаружено, что у крупных форм ослинника 28 хромосом, тогда как у обычных только 14.

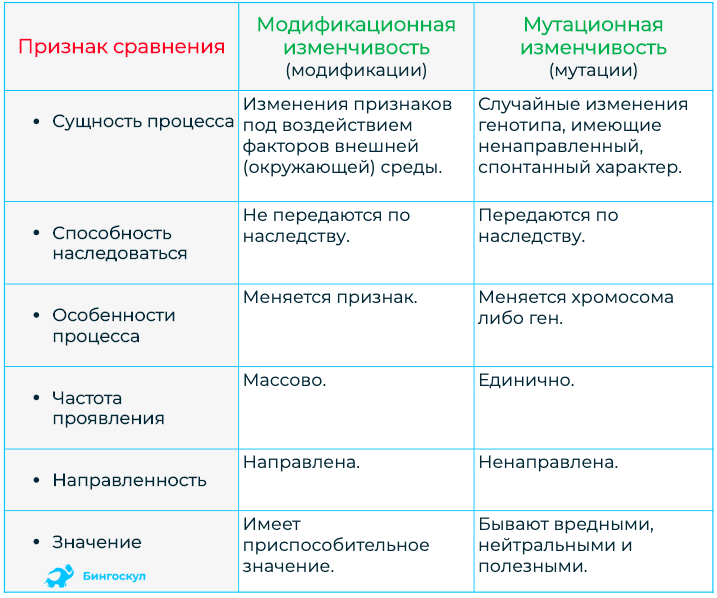
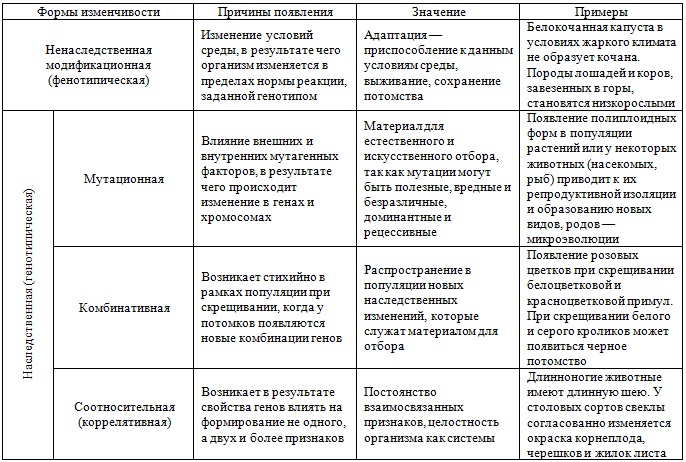
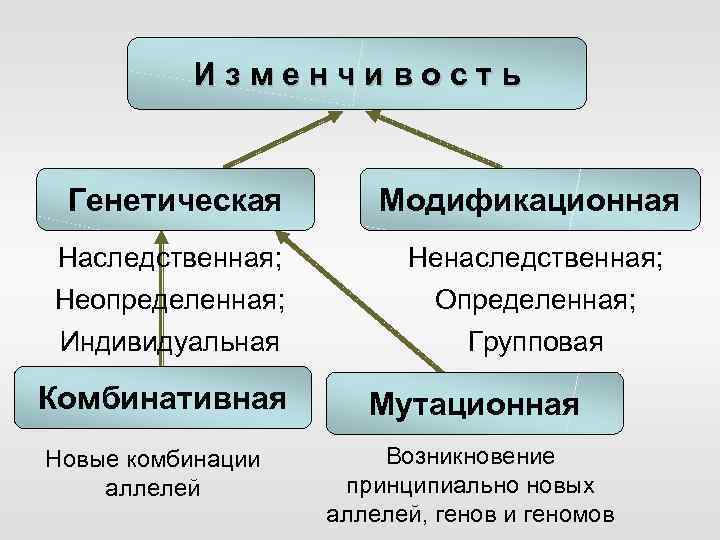




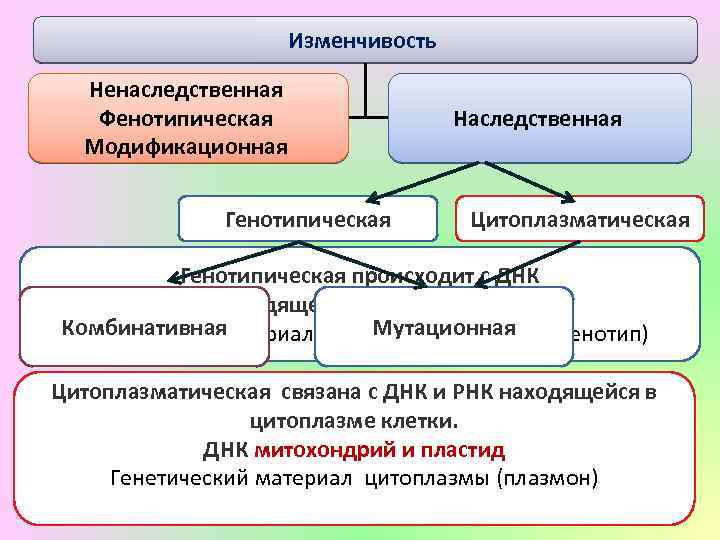
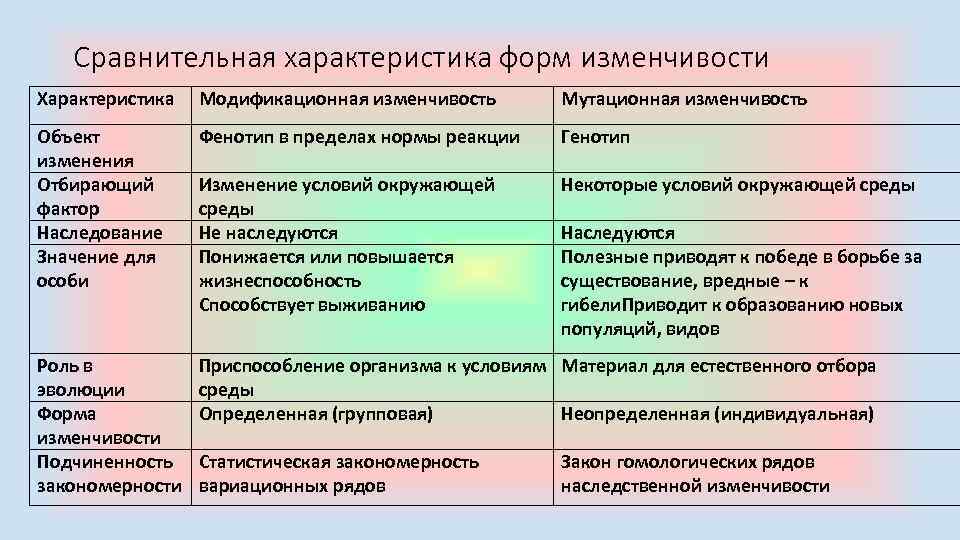
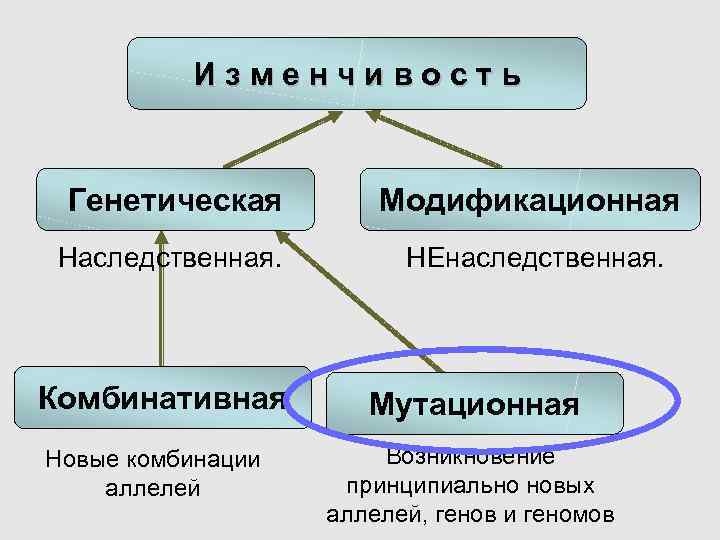
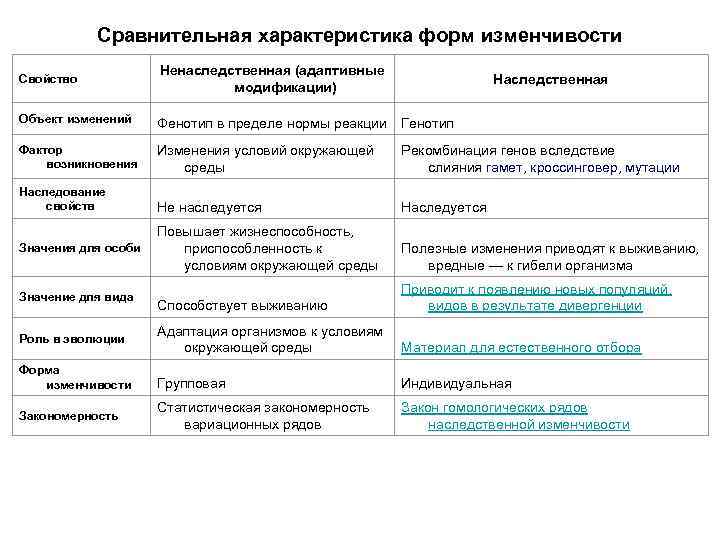
Стало ясно, что мутации – это изменения в генотипе и поэтому являются видом наследственной изменчивости.
Закрепление изменения в потомстве при мутациях – главное отличие от модификационной изменчивости, при которой изменения проявляются только в фенотипе.
Кроме того, в случае мутационной изменчивости значительно расширяется норма реакции, т. е. предел проявления признака. Как следствие, чаще происходят вредные мутации, вызывающие изменения, не совместимые с жизнью.
Мутации также могут быть полезными и нейтральными.
Механизм модификационной изменчивости
Окружающая среда как причина модификаций
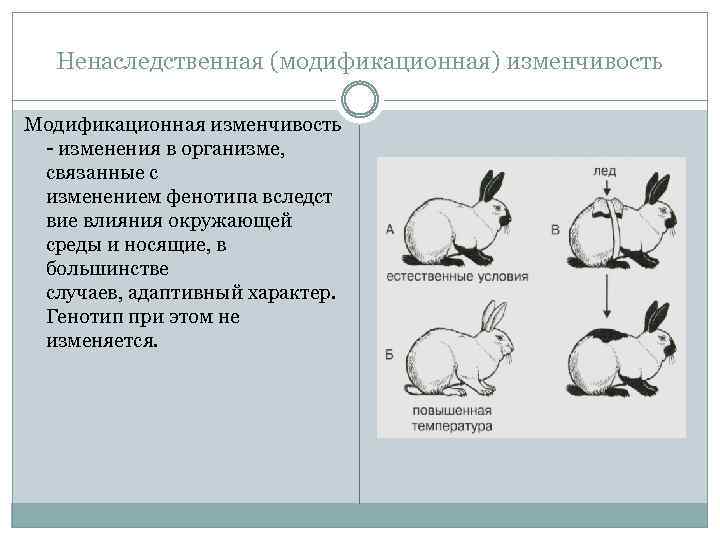
Модификационная изменчивость — это результат не изменений генотипа, а его непосредственной реакции на условия окружающей среды. При модификационной изменчивости наследственный материал не изменяется, — изменяется проявление генов.
Стимулы внешней среды воздействуют на поведение клеток и многоклеточных организмов благодаря наличию чувствительных рецепторов (они имеются не только в органах чувств животных, но и в каждой живой клетке), которые передают цепочки сигналов, меняющих регуляцию функционирования определенных генов. Таким образом, факторы окружающей среды способны регулировать интенсивность выработки клетками специфических белков, от которых зависит развитие, физиология и поведение организма.
Один из примеров — появление «загара» при длительном пребывании человека под лучами ультрафиолета. При воздействии ультрафиолетовых лучей происходит разрушение некоторого количества клеток эпидермиса, что приводит к выделению эндотелина-1 и эйкозаноидов. Они вызывают активацию фермента тирозиназы и его биосинтез. Тирозиназа, в свою очередь, является ключевым ферментом синтеза меланина.
В большинстве случаев модификационная изменчивость способствует адаптации организмов к условиям окружающей среды — генотип реагирует на средовые факторы и происходит перестройка фенотипа в соответствии с изменившимися потребностями организма (например, увеличивается число эритроцитов у человека, поднявшегося в горы). Однако иногда, под влиянием неблагоприятных факторов окружающей среды, например, влиянием тератогенных факторов на беременных, возникают нарушения фенотипа (морфозы), не имеющие какого-либо адаптивного значения. Интересно, что во многих случаях морфозы имеют вид «фенокопий» некоторых известных мутаций, что было пау отмечено в еще в 40-е годы XX века немецким биологом Рихардом Гольдшмидтом.
Норма реакции
Основная статья: Норма реакции
Норма реакции — характерный спектр реакции фенотипа на дозу фактора среды (температуру, влажность, освещенность, обилие корма и пр.). Кривая, описывающая зависимость признака от фактора среды, может быть плавной (с уклоном вверх или вниз), а может быть бимодальной — в этом случае наблюдается явление полифенизма: фенотип представлен двумя или более хорошо различимыми вариантами, не связанных между собой промежуточными вариантами.
Предел проявления модификационной изменчивости организма при неизменном генотипе — норма реакции. Норма реакции обусловлена генотипом и различается у разных особей данного вида. Фактически норма реакции — спектр возможных уровней экспрессии генов, из которого выбирается уровень экспрессии, наиболее подходящий для данных условий окружающей среды. Усиленное кормление приведёт к увеличению массы животного, однако она будет находиться в пределах нормы реакции, характерной для данного вида или породы. Норма реакции генетически детерминирована и наследуется. Для разных признаков пределы нормы реакции сильно различаются. Например, широкие пределы нормы реакции имеют величина удоя, продуктивность злаков и многие другие количественные признаки, узкие пределы — интенсивность окраски большинства животных, температура тела у теплокровных, число позвонков и пальцев у позвоночных.
Качественные признаки (наличие или отсутствие определенных органов и частей тела, паттерны рисунка на шкуре — пятна, полоски и пр., стадии жизненного цикла) проявляют фенотипическую пластичность гораздо реже, чем количественные. Но такие примеры все же встречаются. К примеру, у тлей в зависимости от количества питания и плотности населения, могут формироваться бескрылые или крылатые особи, также они могут переключаться между бесполым и половым размножением, откладывать яйца или переходить к живорождению. Следует отметить, что граница между количественными и качественными признаками иногда весьма условна.




Саранча встречается в двух основных формах: одиночной (вверху) и стадной (внизу), хорошо различимых морфологически, а также различающихся по поведению и физиологии. Переход между формами зависит от плотности популяции и может занимать одно-несколько поколений.
Генетика
Генетика популяций — наука о генетической структуре природных популяций и генетических процессах, происходящих в ней, таких, как дрейф генов, миграция, мутация и отбор.
Все организмы состоят из больших популяций, в которых по законам генетики поддерживается равновесие генетического материала. Однако это равновесие постоянно нарушается мутационными процессами, миграциями, дрейфом генов и другими факторами.
Все разнообразие в человеческих популяциях — есть результат мутационных изменений. Видный генетик С. С. Четвериков (1882—1959) внес существенный вклад в доказательство связи генетики с эволюцией. Он показал, что первые элементарные процессы начинаются в популяциях. Природные популяции при относительной фенотипичной однородности по генетической структуре разнородны и насыщены множеством открытых мутаций, образующих резерв {генетический груз) наследственной изменчивости.
Под генетической структурой понимают соотношение в ней разных генотипов и аллельных генов. Английский математик Харди и немецкий врач Вайнберг установили, что при идеальных условиях — крупной популяции отсутствии мутаций, миграций и отбора — соотношение генотипов и аллельных генов во всех поколениях постоянно.
Резерв наследственной изменчивости в популяции образуется за счет мутации. Доминантные мутации возникают редко, проявляются сразу и подвергаются отбору,
Рецессивные мутации у гетерозиготных организмов фенотипически не проявляются, но при скрещивании насыщают генофонд популяции и образуют новые генотипы.
Генофонд популяций пополняется также за счет генного потока — миграции особей из других популяций, приносящих новые гены. Они, также, как мутации, при скрещиваниях первое время у гетерозиготных организмов не проявляются. Одним из путей относительно быстрого изменения частот генов является случайное распределение генов, называемое дрейфом генов.
Дрейф генов, случайная, нецеленаправленная смена частоты встречаемости аллелей в популяции, обуславливаемая периодическими популяционными волнами. Чаще всего дрейф генов встречается в малочисленных популяциях. В результате дрейфа генов в популяции может возрастать частота встречаемости редких аллелей, некоторые аллели могут исчезать; длительный период могут сохраняться мутантные аллели, что снижает приспособленность особей к условиям жизни.
Резерв наследственной информации образуется еще и за счет комбинативной изменчивости, при которой в одном генотипе объединяются и обезвреживаются разнонаправленные мутации.
Накапливаясь в популяции, скрытые мутации частично переходят в гомозиготное состояние и тогда проявляются фенотипически. В постоянных условиях стабилизирующий отбор (отбор в пользу нормы признака) устраняет их как несоответствующие условиям среды.




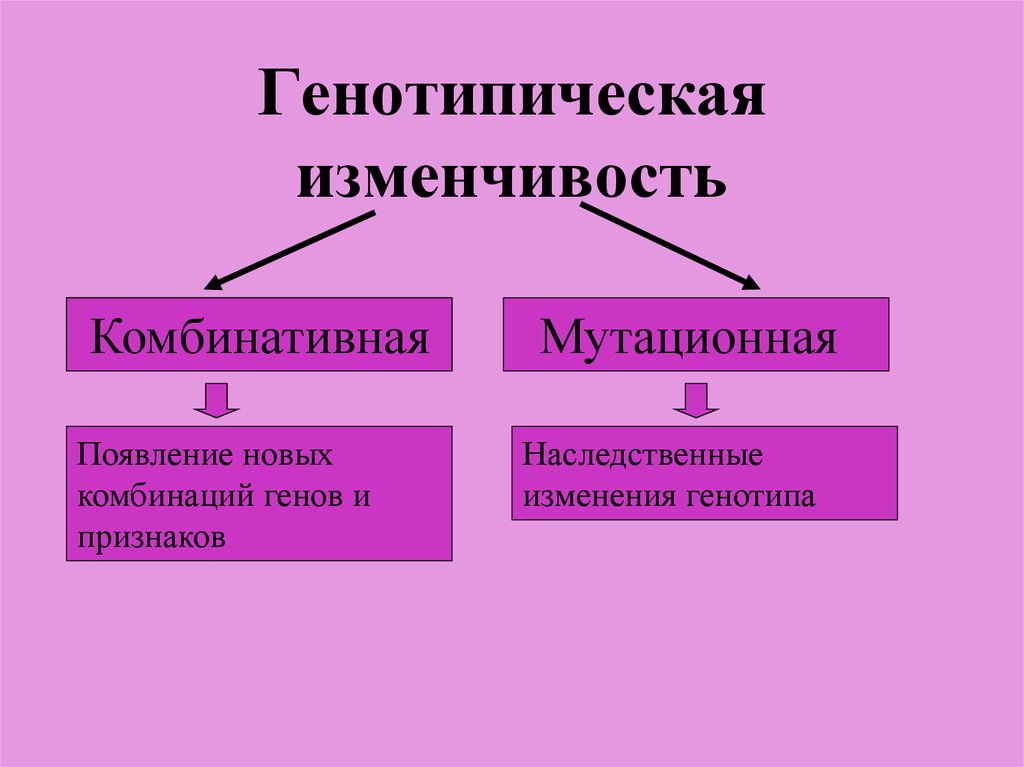
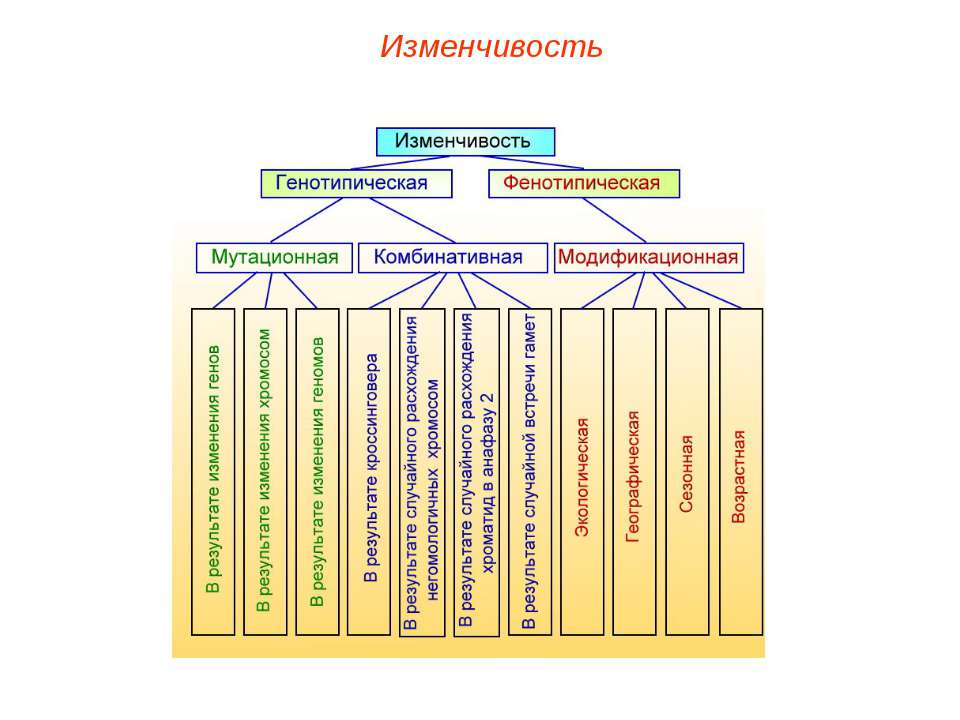



В меняющихся условиях, при действии движущего отбора (отбора некоторых отклонений от установившейся нормы признаков), резерв наследственной изменчивости позволяет популяции приспосабливаться к новым условиям среды. Чем больше генотипов в популяции, тем шире ее норма реакции, тем вероятнее ее выживание в меняющихся условиях и возможность полнее использовать новые места обитания.
Каждый биологический вид обладает уникальным генофондом, поэтому одной из важнейших задач человечества является охрана генофонда естественных популяций организмов.
Классификация мутаций
По причинам, вызвавшим мутации, их подразделяют на спонтанные и индуцированные. Спонтанные (самопроизвольные)мутации происходят под действием естественных мутагенных факторов внешней среды без вмешательства человека, например наследственные болезни обмена веществ. Их причинами являются ошибки репликации и репарации ДНК, действие перекисей и альдегидов, образующихся в организме, различные виды естественных излучений. Индуцированные мутации — результат направленного воздействия определенных мутагенных факторов. Впервые они были получены в 1925 г. Г.А. Надсоном и Г.С. Филипповым при облучении грибов радием.
По мутировавшим клеткам мутации подразделяют на гаметические и соматические. Гаметические мутации (генеративные) происходят в половых клетках, проявляются у потомков, передаются по наследству при половом размножении (гемофилия, фенилкетонурия). Соматические мутации происходят в соматических клетках, передаются по наследству только при вегетативном размножении и проявляются у самой особи (разный цвет глаз у одного человека, белая прядь волос, опухоли).
По исходу для организма мутации бывают: отрицательные — летальные, несовместимые с жизнью (например, отсутствие головного мозга) и полулетальные — снижающие жизнеспособность организма (например, болезнь Дауна); нейтральные — существенно не влияющие на процессы жизнедеятельности (например, веснушки); положительные — повышающие жизнеспособность (например, появление четырехкамерного сердца в процессе эволюции хордовых животных).
Роль в эволюции
На наследственной изменчивости основано всё разнообразие индивидуальных различий, которые включают:
- Как резкие качественные различия, не связанные друг с другом переходными формами, так и чисто количественные различия, образующие непрерывные ряды, в которых близкие члены ряда могут отличаться друг от друга сколь угодно мало;
- Как изменения отдельных признаков и свойств (независимая изменчивость), так и взаимосвязанные изменения ряда признаков (коррелятивная изменчивость);
- Как изменения, имеющие приспособительное значение (адаптивная изменчивость), так и изменения «безразличные» или даже снижающие жизнеспособность их носителей (неадаптивная изменчивость).
Все эти типы наследственных изменений составляют материал эволюционного процесса (см. Микроэволюция).
В индивидуальном развитии организма проявление наследственных признаков и свойств всегда определяется не только основными, ответственными за данные признаки и свойства генами, но и их взаимодействием со многими другими генами, составляющими генотип особи, а также условиями внешней среды, в которой протекает развитие организма.
Неоспоримо важна точность при передаче генетической информации в ряду поколений, однако чрезмерная консервация генетической информации, заключенной в отдельных генетических локусах, может быть вредной для организма и вида в целом.
Эволюционно сложившиеся отношения между точностью функционирования генетических систем и частотой ошибок, возникающих при воспроизведении генетической информации отдельных генетических локусов, четко сбалансированы между собой, и уже установлено, что в ряде случаев являются регулируемыми. Запрограммированные и случайные наследуемые изменения генома, называемые мутациями, могут сопровождаться колоссальными количественными и качественными изменениями в экспрессии генов.
Длительная модификационная изменчивость
В большинстве случаев модификационная изменчивость носит ненаследственный характер и является лишь реакцией генотипа данной особи на условия среды с последующим изменением фенотипа. Однако известны и примеры наследуемых средозависимых изменений, описанные у некоторых бактерий, простейших и многоклеточных эукариот. Чаще всего в настоящее время эти случаи определяют как «трансгенерационное эпигенетическое наследование», но в учебниках советского времени к таким случаям применяют понятие «длительная модификация».
Для понимания возможного механизма наследования модификационной изменчивости рассмотрим сначала понятие генетического триггера.
Например, в оперонах бактерий содержатся, кроме структурных генов, два участка — промотор и оператор. Оператор некоторых оперонов находится между промотором и структурными генами (у других он входит в состав промотора). Если оператор связан с белком, который называется репрессором, то вместе они не дают двигаться РНК-полимеразе по цепи ДНК. У бактерий E. сoli можно наблюдать подобный механизм. При недостатке лактозы и избытке глюкозы вырабатывается белок-репрессор (Lacl), который присоединяется к оператору, не давая РНК-полимеразе синтезировать мРНК для трансляции фермента, который расщепляет лактозу. Однако при попадании лактозы в цитоплазму бактерии лактоза (вещество-индуктор) присоединяется к белку-репрессору, изменяя его конформацию, что приводит к диссоциации репрессора от оператора. Это обуславливает начало синтеза фермента для расщепления лактозы.









У бактерий при делении вещество-индуктор (в случае с E. coli — лактоза) передаётся в цитоплазму дочерней клетки и запускает диссоциацию белка-репрессора от оператора, что влечет за собой проявление активности фермента (лактазы) для расщепления лактозы у палочек даже при отсутствии этого дисахарида в среде.
Если оперона два и если они взаимосвязаны (структурный ген первого оперона кодирует белок-репрессор для второго оперона и наоборот), они образуют систему, которая называется триггером. При активном состоянии первого оперона отключен второй. Однако под действием окружающей среды может быть заблокирован синтез белка-репрессора первым опероном, и тогда происходит переключение триггера: активным становится второй оперон. Такое состояние триггера может наследоваться следующими поколениями бактерий. Молекулярные триггеры могут обеспечивать длительные модификации и у одноклеточных эукариот (в частности, подобные явления отмечались у дрожжей, инфузорий и др.).
Наиболее любопытны и наименее понятны с точки зрения механизма случаи наследования индуцированных средой изменений у многоклеточных организмов — животных и растений. Прогресс в области эпигенетики, заявившей свои права в течение последних двух десятилетий, позволяет находить естественные объяснения подобным явлениям там, где раньше они казались невозможными. Наиболее вероятным фактором, который позволяет передавать «благоприобретенные» признаки от родительской особи к потомству видится малая некодирующая РНК. Эти молекулы могут попадать в зиготу (оплодотворенную яйцеклетку) с цитоплазмой и содержимым ядер яйцеклетки и сперматозоида и управлять включениями и выключениями генов развивающегося организма, влияя на формирующийся фенотип.
Этапы селекционной работы
I этап селекционной работы
Исходное сортовое и видовое разнообразие растений и животных — объекты селекционной работы (без знания исходного материала, без изучения его происхождения и эволюции невозможно совершенствование существующих форм животных и растений).
На данном этапе используются работы Н. И. Вавилова по установлению центров происхождения культурных растений в очагах древнейшего земледелия, созданию их коллекции и использованию в качестве исходного материала. Таких центров восемь.
- Южноазиатский тропический центр. Тропическая Индия, Индокитай, Южный Китай, острова Юго-Восточной Азии. Исключительно богат культурными растениями (около ½ известных видов культурных растений). Родина риса, сахарного тростника, множества плодовых и овощных культур.
- Восточноазиатский центр. Центральными Восточный Китай, Япония, остров Тайвань, Корея. Родина сои, нескольких видов проса, множества плодовых и овощных культур. Этот центр тоже богат видами культурных растений — около 20% мирового многообразия.
- Юго-Западноазиатский центр. Малая Азия, Средняя Азия, Иран, Афганистан, Северо-Западная Индия. Родина нескольких форм пшеницы, ржи, многих зерновых, бобовых, винограда, плодовых. В нем возникло 14% мировой культурной флоры.
- Средиземноморский центр. Страны, расположенные но берегам Средиземного моря. Этот центр, где располагались величайшие древние цивилизации, дал около 11% видов культурных растений. В их числе маслины, многие кормовые растения (клевер, одноцветковая чечевица), многие овощные (капуста) и кормовые культуры.
- Абиссинский центр. Небольшой район Африканского материка с очень своеобразной флорой культурных растений. Очевидно, очень древний очаг самобытной земледельческой культуры. Родина зернового сорго, одного вида бананов, масличного растения нута, ряда особых форм пшеницы и ячменя.
- Центральноамериканский центр. Южная Мексика. Родина кукурузы, длинноволокнистого хлопчатника, какао, ряда тыквенных, фасоли — всего около 90 видов культурных растений.
- Андийский (Южноамериканский) центр. Включает часть районов Андийского горного хребта вдоль западного побережья Южной Америки. Родина многих клубненосных растений, в том числе картофеля, некоторых лекарственных растений (кокаиновый куст, хинное дерево и др.)
Подавляющее большинство культурных растений связано в своем происхождении с одним или несколькими из перечисленных выше географических центров.
II этап — скрещивание (гибридизация)
Бывает двух видов:
- Близкородственное — инбридинг (позволяющее перевести рецессивные гены в гомозиготное состояние);
- Неродственное (помогающее объединить в одном организме ценные признаки разных форм).
III этап — отбор — завершающий этап селекции.
Известно две формы отбора:
- массовый (выделение группы особей, сходных по фенотипу, но дающих расщепление при размножении)
- индивидуальный (выделение единичных ценных форм и раздельное выращивание Потомства каждой особи) приводит к созданию сорта или породы чистой линии.
В селекции растений широко используется инбридинг, полиплоидия, искусственный мутагенез, отдаленная гибридизация.
В области селекции растений много сделали известные селекционеры-генетики: И. В. Мичурин и Г. Д. Карнеченко, II. В. Цицин, П. II. Лукьяненко, В. Н. Ремесло, В. С. Пустовойт и л р.
Ими были выведены высокоурожайные сорта сахарной свеклы, гречихи, хлопчатника; высокопродуктивные кубанские сорта пшеницы, украинские сорта «Мироновская-808, «Юбилейная-50», «Харьковская-63» и др.
Селекция животных отличается от таковой у растении: животные дают мало потомков, у них позднее наступает половозрелость, они не размножаются вегетативно, отсутствует самооплодотворение.
В селекции животных используют гибридизацию и отбор (массовый и индивидуальный), инбридинг и другие методы (М. Ф. Иванов, Н. С, Батурин и др.)




Селекция микроорганизмов — молодая, развивающаяся отрасль селекции. Ее задача — получение высокопродуктивных микроорганизмов путем воздействия на исходные формы лучами Рентгена, ультрафиолетовыми лучами и химическими мутагенами.
Чередование обработки мутагенами с отбором позволяет выделять штаммы, по продуктивности в десятки раз превосходящие исходный.