Нарушения, связанные с процессами рекомбинации
Поскольку рекомбинация требует физического связывания в хромосомах, это важный шаг в правильной сегрегации во время мейоза. Если адекватная рекомбинация не происходит, результатом может быть значительная патология.
Нерасщепление хромосом или ошибки в сегрегации является одной из наиболее частых причин абортов и аномалий хромосомного происхождения, таких как трисомия 21 хромосомы, которая вызывает синдром Дауна..
Хотя рекомбинация обычно является довольно точным процессом, участки генома, которые повторяются, и гены, имеющие несколько копий вдоль генома, являются элементами, склонными к неравное пересечение.
Это скрещивание вызывает различные клинические признаки, включая частые заболевания, такие как талассемия и аутизм..
Митоз и негомологичные хромосомы
В эукариотических клетках скрещивание также может происходить во время митоза. При этом получаются две клетки с идентичным генетическим материалом. Любой кроссовер, который происходит между гомологичными хромосомами в митозе, не производит новую комбинацию генов.
Пересечение, которое происходит в негомологичных хромосомах, может произвести мутацию, известную как транслокация. Она происходит, когда сегмент хромосомы отделяется от и перемещается в новое положение на негомологичной молекуле. Этот тип мутации может быть опасен, так как часто приводит к развитию раковых клеток.

Что такое гомологичная рекомбинация?
Гомологичная рекомбинация — это клеточный феномен, который включает физический обмен последовательностями ДНК между двумя хромосомами. Рекомбинация включает в себя набор генов, известных как гены запись. Эти коды для различных ферментов, участвующих в процессе.
Молекулы ДНК считаются «гомологичными», когда они имеют сходные или идентичные последовательности из более чем 100 пар оснований. ДНК имеет небольшие области, которые могут отличаться друг от друга, и эти варианты известны как аллели.
У живых существ вся ДНК считается рекомбинантной ДНК. Обмен генетическим материалом между хромосомами происходит непрерывно, смешивая и переставляя гены в хромосомах..
Этот процесс происходит, очевидно, в мейозе. Конкретно в фазе, где хромосомы спарены в первом клеточном делении. На этом этапе происходит обмен генетическим материалом между хромосомами..
Исторически этот процесс обозначен в литературе с использованием англосаксонского слова пересечение. Это событие является одним из результатов гомологичной рекомбинации.
Частота пересечение между двумя генами одной и той же хромосомы зависит в основном от расстояния между ними; чем меньше физическое расстояние между ними, тем ниже частота обмена.
Связь между генерацией и рекомбинацией

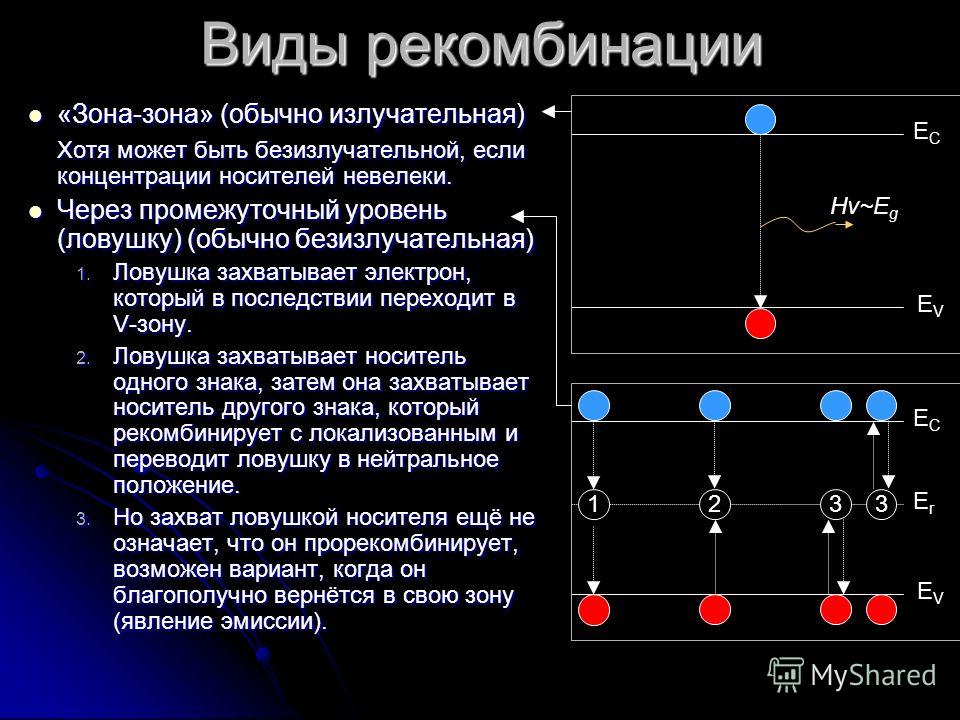
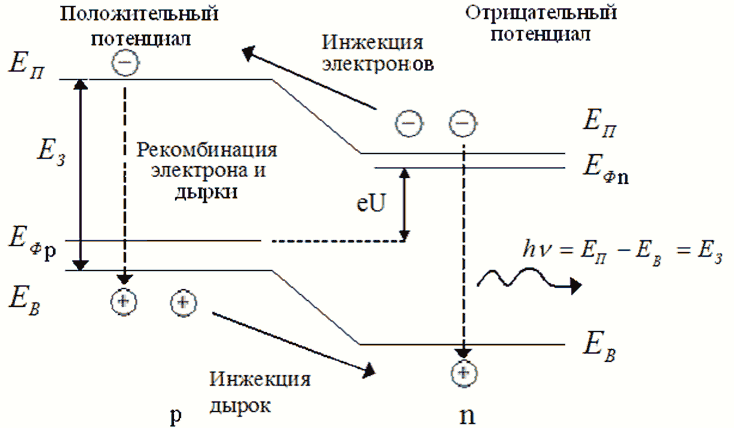
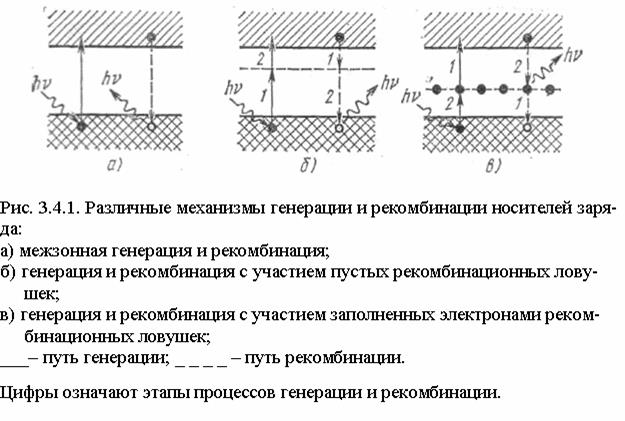
На следующем изображении показано изменение генерируемых избыточных носителей (зеленый: электроны и фиолетовый: дырки) при увеличении интенсивности света (скорость генерации / см ) в центре собственной полупроводниковой планки. Электроны имеют более высокую постоянную диффузии, чем дырки, что приводит к меньшему количеству избыточных электронов в центре по сравнению с дырками.3{\ displaystyle ^ {3}}
Рекомбинация и генерация всегда происходят в полупроводниках, как оптически, так и термически. Как предсказывает термодинамика , материал, находящийся в тепловом равновесии, будет иметь скорости генерации и рекомбинации, которые уравновешены, так что чистая плотность носителей заряда остается постоянной. Результирующая вероятность заполнения энергетических состояний в каждой энергетической зоне определяется статистикой Ферми – Дирака .




Произведение плотностей электронов и дырок ( и ) является константой в состоянии равновесия, поддерживаемой рекомбинацией и генерацией, происходящей с равными скоростями. Когда есть избыток носителей (т. Е. ), Скорость рекомбинации становится больше, чем скорость генерации, возвращая систему к равновесию. Аналогичным образом, когда существует дефицит носителей (т. Е. ), Скорость генерации становится больше, чем скорость рекомбинации, снова возвращая систему к равновесию. Когда электрон перемещается из одной энергетической зоны в другую, энергия и импульс, которые он потерял или приобрел, должны идти или исходить от других частиц, участвующих в процессе (например, фотонов , электронов или системы колеблющихся атомов решетки ).
п{\ displaystyle n}п{\ displaystyle p}(попознак равнопя2){\ Displaystyle (п_ {о} р_ {о} = п_ {я} ^ {2})}пп>пя2{\ displaystyle np> n_ {i} ^ {2}}пп<пя2{\ displaystyle np <n_ {i} ^ {2}}
Излучательная рекомбинация
Межзонная излучательная рекомбинация
Межзонная рекомбинация — это название процесса перехода электронов из зоны проводимости в валентную зону излучательным способом. Во время межзонной рекомбинации, формы спонтанного излучения , энергия, поглощенная материалом, высвобождается в виде фотонов . Обычно эти фотоны содержат такую же или меньшую энергию, чем первоначально поглощенные. Этот эффект — это то, как светодиоды создают свет. Поскольку фотон несет относительно небольшой импульс , излучательная рекомбинация имеет значение только в материалах с прямой запрещенной зоной . Этот процесс также известен как бимолекулярная рекомбинация .
Этот тип рекомбинации зависит от плотности электронов и дырок в возбужденном состоянии, обозначенной и соответственно. Представим скорость генерации носителей как, а излучательную рекомбинацию как . Затем, рассматривая случай, когда происходит только межполосная рекомбинация, мы можем выразить изменение плотности носителей как функцию времени как:п(т){\ Displaystyle п (т)}п(т){\ displaystyle p (t)}грамм{\ displaystyle G}рр{\ displaystyle R_ {r}}
В тепловом равновесии излучательная рекомбинация равна скорости тепловой генерации , с законом действия масс ее можно записать как:р{\ displaystyle R_ {0}}грамм{\ displaystyle G_ {0}} ппзнак равнопя2{\ displaystyle n_ {0} p_ {0} = n_ {i} ^ {2}}
где называется скоростью излучательной рекомбинации, — собственная плотность носителей и — равновесные плотности носителей. При наличии электронно-дырочных пар зарядовая нейтральность сохраняется, а неравновесные плотности определяются выражением:Bр{\ displaystyle B_ {r}}пя{\ displaystyle n_ {i}}п{\ displaystyle n_ {0}}п{\ displaystyle p_ {0}}Δпзнак равноΔп{\ displaystyle \ Delta n = \ Delta p}
Тогда чистая скорость рекомбинации становится,рр{\ displaystyle R_ {r}}
Решая это уравнение с учетом нейтральности заряда, радиационное время жизни определяется выражениемτр{\ displaystyle \ tau _ {r}}
Вынужденное излучение
Вынужденное излучение — это процесс, в котором падающий фотон взаимодействует с возбужденным электроном, заставляя его рекомбинировать и излучать фотон с теми же свойствами, что и падающий, с точки зрения фазы , частоты , поляризации и направления движения. Вынужденное излучение вместе с принципом инверсии населенностей лежат в основе работы лазеров и мазеров . Эйнштейн в начале двадцатого века показал, что если возбужденный и основной уровни невырождены, то скорость поглощения и скорость вынужденного излучения одинаковы. Иначе, если уровень 1 и уровень 2 являются -кратно вырожденными и -кратно вырожденными соответственно, новое отношение будет следующим:W12{\ displaystyle W_ {12}}W21 год{\ displaystyle W_ {21}}грамм1{\ displaystyle g_ {1}}грамм2{\ displaystyle g_ {2}}
Значение
Отслеживание движения генов в результате кроссоверов оказалось весьма полезным для генетиков. Это дает возможность определить, насколько далеки друг от друга два гена в хромосоме. Наука также может использовать этот метод для вывода о наличии определенных генов. Одна молекула в связанной паре служит маркером для выявления наличия другой. Это используется, чтобы обнаружить присутствие болезнетворных генов.
Частота рекомбинации между двумя наблюдаемыми локусами является значением пересечения. Она зависит от взаимного расстояния наблюдаемых генетических очагов. Для любого фиксированного набора условий окружающей среды рекомбинация в определенной области структуры связей (хромосомы) имеет тенденцию быть постоянной. То же самое справедливо для значения пересечения, которое используется при создании генетических карт.

Классификация: тирозин- против серин-рекомбиназы
 Рис. 1. Тир-рекомбиназы: детали стадии кроссовера. Вверху: традиционный вид, включающий обмен нитями с последующим переносом веток (корректура). Механизм происходит в рамках синаптического комплекса (1), включающего оба сайта ДНК в параллельной ориентации. В то время как ветвящаяся миграция напрямую объясняет специфические требования гомологии и обратимость процесса, она не может быть согласована с движениями субъединиц рекомбиназы в трех измерениях. Внизу: текущий вид. Две одновременные замены цепей, каждая из которых зависит от комплементарности трех последовательных оснований на краях спейсера из 8 пар оснований (или рядом с ними) (пунктирные линии указывают на спаривание оснований). Дидактические сложности возникают из-за того, что в этой модели синаптический комплекс должен вмещать оба субстрата в антипараллельной ориентации. Этот синаптический комплекс (1) возникает в результате ассоциации двух индивидуальных субъединиц рекомбиназы («протомеры»; серые овалы) с соответствующим сайтом-мишенью. Его образование зависит от межпротомерных контактов и изгиба ДНК, которые, в свою очередь, определяют субъединицы (зеленые), играющие активную роль во время первой реакции кроссовера. Оба изображения иллюстрируют только половину соответствующего пути. Эти части разделяются стадией соединения Холлидея / изомеризации перед высвобождением продукта (3).
Рис. 1. Тир-рекомбиназы: детали стадии кроссовера. Вверху: традиционный вид, включающий обмен нитями с последующим переносом веток (корректура). Механизм происходит в рамках синаптического комплекса (1), включающего оба сайта ДНК в параллельной ориентации. В то время как ветвящаяся миграция напрямую объясняет специфические требования гомологии и обратимость процесса, она не может быть согласована с движениями субъединиц рекомбиназы в трех измерениях. Внизу: текущий вид. Две одновременные замены цепей, каждая из которых зависит от комплементарности трех последовательных оснований на краях спейсера из 8 пар оснований (или рядом с ними) (пунктирные линии указывают на спаривание оснований). Дидактические сложности возникают из-за того, что в этой модели синаптический комплекс должен вмещать оба субстрата в антипараллельной ориентации. Этот синаптический комплекс (1) возникает в результате ассоциации двух индивидуальных субъединиц рекомбиназы («протомеры»; серые овалы) с соответствующим сайтом-мишенью. Его образование зависит от межпротомерных контактов и изгиба ДНК, которые, в свою очередь, определяют субъединицы (зеленые), играющие активную роль во время первой реакции кроссовера. Оба изображения иллюстрируют только половину соответствующего пути. Эти части разделяются стадией соединения Холлидея / изомеризации перед высвобождением продукта (3).
 Рис. 2. Сер-рекомбиназы: (по существу необратимый) путь вращения субъединиц. В отличие от Tyr-рекомбиназ, четыре участвующих нити ДНК разрезаются синхронно в точках, разнесенных только на 2 п.н. (оставляя мало места для проверки). Вращение субъединицы (180 °) позволяет обмениваться цепями, будучи ковалентно связанными с белком-партнером. Промежуточное воздействие двухцепочечных разрывов несет риск запуска незаконной рекомбинации и, следовательно, вторичных реакций. Здесь синаптический комплекс возникает в результате ассоциации предварительно сформированных димеров рекомбиназы с соответствующими сайтами-мишенями (CTD / NTD, C- / N-концевой домен). Как и в случае Tyr-рекомбиназ, каждый сайт содержит два плеча, в каждом из которых находится один протомер. Поскольку оба плеча структурированы немного по-разному, субъединицы становятся конформационно настроенными и, таким образом, готовятся к их соответствующей роли в цикле рекомбинации. В отличие от представителей Tyr-класса, путь рекомбинации преобразует два разных сайта субстрата (attP и attB) в сайт-гибриды (attL и attR) . Это объясняет необратимую природу этого конкретного пути рекомбинации, который может быть преодолен только с помощью вспомогательных «факторов направленности рекомбинации» (RDF).
Рис. 2. Сер-рекомбиназы: (по существу необратимый) путь вращения субъединиц. В отличие от Tyr-рекомбиназ, четыре участвующих нити ДНК разрезаются синхронно в точках, разнесенных только на 2 п.н. (оставляя мало места для проверки). Вращение субъединицы (180 °) позволяет обмениваться цепями, будучи ковалентно связанными с белком-партнером. Промежуточное воздействие двухцепочечных разрывов несет риск запуска незаконной рекомбинации и, следовательно, вторичных реакций. Здесь синаптический комплекс возникает в результате ассоциации предварительно сформированных димеров рекомбиназы с соответствующими сайтами-мишенями (CTD / NTD, C- / N-концевой домен). Как и в случае Tyr-рекомбиназ, каждый сайт содержит два плеча, в каждом из которых находится один протомер. Поскольку оба плеча структурированы немного по-разному, субъединицы становятся конформационно настроенными и, таким образом, готовятся к их соответствующей роли в цикле рекомбинации. В отличие от представителей Tyr-класса, путь рекомбинации преобразует два разных сайта субстрата (attP и attB) в сайт-гибриды (attL и attR) . Это объясняет необратимую природу этого конкретного пути рекомбинации, который может быть преодолен только с помощью вспомогательных «факторов направленности рекомбинации» (RDF).
На основе гомологии аминокислотной последовательности и механистического родства большинство сайт-специфичных рекомбиназ сгруппированы в одно из двух семейств: семейство тирозиновых (Tyr) рекомбиназ или семейство серин (Ser) рекомбиназ . Названия происходят от консервативного нуклеофильного аминокислотного остатка, присутствующего в каждом классе рекомбиназы, который используется для атаки ДНК и который становится ковалентно связанным с ней во время обмена цепью. Самые ранние идентифицированные члены семейства сериновых рекомбиназ были известны как резольвазы или ДНК-инвертазы , в то время как член-основатель тирозиновых рекомбиназ, лямбда- фаговая интеграза (с использованием сайтов узнавания attP / B), отличается от хорошо известных сейчас ферментов, таких как Cre ( из фага P1 ) и FLP (из дрожжей Saccharomyces cerevisiae ). Известные сериновые рекомбиназы включают ферменты, такие как гамма-дельта-резольваза (из транспозона Tn 1000 ), резольваза Tn3 (из транспозона Tn3) и интеграза φ C31 (из фага φ C31).
Хотя отдельные члены двух семейств рекомбиназ могут выполнять реакции с одинаковыми практическими результатами, семейства не связаны друг с другом, имея разные белковые структуры и механизмы реакции. В отличие от тирозиновых рекомбиназ, сериновые рекомбиназы имеют высокую модульность, на что сначала намекали биохимические исследования, а затем показали кристаллографические структуры. Знание этих белковых структур может оказаться полезным при попытке реконструировать рекомбиназные белки в качестве инструментов для генетических манипуляций.
Рекомбинация (биология)
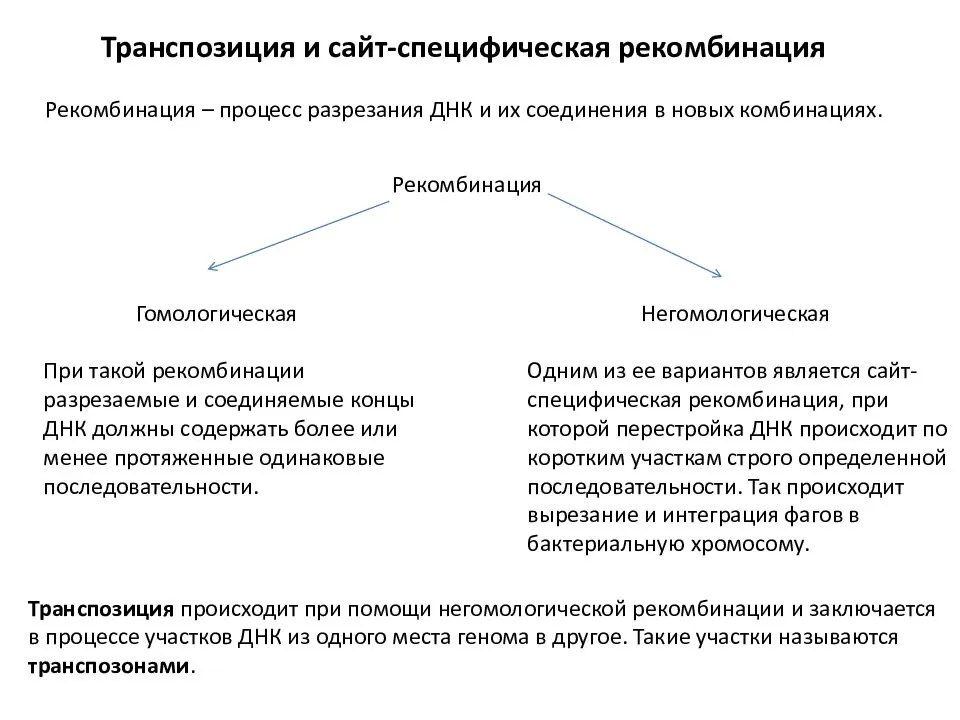
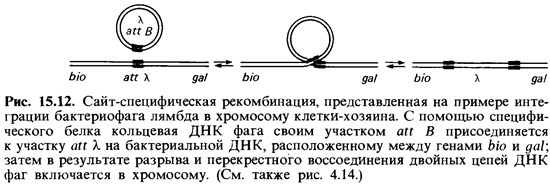
Подробное ее описание будет дано ниже, а пока остановимся кратко на двух других типах рекомбинации, происходящей под контролем ферментов, опознающих специфические последовательности нуклеотидов, присутствующие на одной или двух рекомбинирующих молекулах. С помощью этого типа рекомбинации бактериальные вирусы и мобильные элементы перемещаются по геному.




Дата добавления: 2015-05-10; просмотров: 775; Опубликованный материал нарушает авторские права? | Защита персональных данных |
Не нашли то, что искали? Воспользуйтесь поиском:
, перераспределение (перекомбинирование) генетического материала родителей, в результате чего у потомков появляются новые сочетания генов, определяющие новые сочетания признаков. Другими словами, сочетание признаков у потомков никогда не повторяет сочетания признаков ни одного из родителей. Рекомбинация – основа комбинативной изменчивости, обеспечивающей бесконечное разнообразие особей внутри вида и неповторимость каждой из них.
ссылки
- Альбертс Б., Джонсон А.Д., Льюис Дж., Морган Д., Рафф М., Робертс К., Уолтер П. (2014) Молекулярная биология клетки (6-е издание). W. W. Norton & Company, Нью-Йорк, Нью-Йорк, США.
- Белл, J.C., Kowalczykowski, S.C. (2016) Механика и одно-молекулярный опрос рекомбинации ДНК. Ежегодный обзор биохимии, 85: 193-226.
- Прадо, Ф. () Гомологичная рекомбинация: к форку и дальше. Genes, doi: 10.3390 / genes9120603
- Griffiths A.J.F., Wessler R., Carroll S.B., Doebley J. (2015). Введение в генетический анализ (11-е изд.). Нью-Йорк: У. Х. Фриман, Нью-Йорк, Нью-Йорк, США.
- Tock, A.J., Henderson, I.R. (2018) Горячие точки для инициации мейотической рекомбинации. Границы в области генетики, дои: 10.3389 / fgene.2018.00521
- Валь, А., Баттести, А., Ансальди, М. (2018) Salmonella enterica: движущая сила в изменении генома и физиологии их бактериального хозяина? Молекулярная микробиология, doi: 10,1111 / mmi.14167.
- Wright W. D., Shah S., Heyer, W. D. (2018) Гомологичная рекомбинация и репарация ДНК двухцепочечных разрывов. Журнал биологической химии, 293: 10524-10535
Значение слова РЕКОМБИНАЦИЯ в Энциклопедии Биология
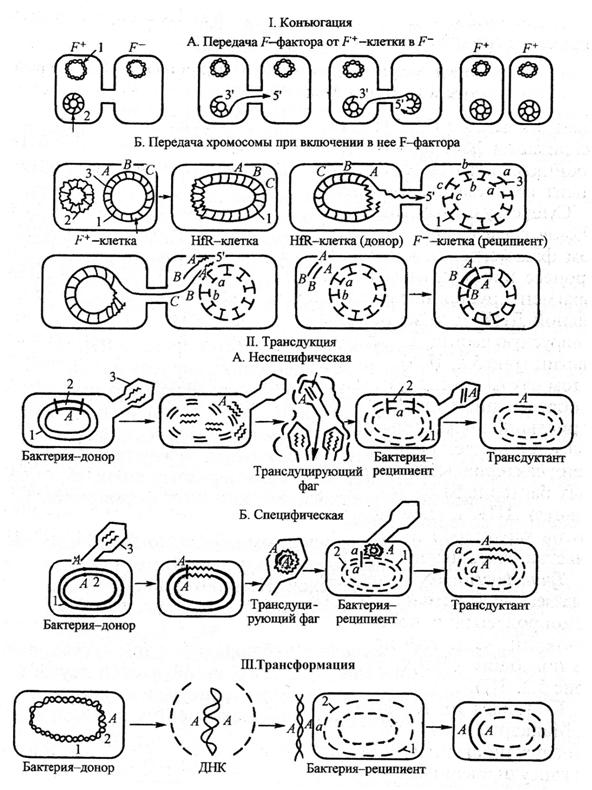
В процессе репликации фага внутри бактерий фрагмент бактериальной ДНК проникает в фаговую частицу и переносится в реципиентную бактерию во время фаговой инфекции. Существует три типа трансдукции:
общая трансдукция (или неспецифическая) – перенос бактериофагом фрагмента любой части бактериальной хромосомы – происходит вследствие того, что бактериальная ДНК фрагментируется после фаговой инфекции и кусочек бактериальной ДНК того же размера, что и фаговая ДНК, проникает в вирусную, формируя дефектную фаговую частицу с частотой приблизительно 1 на 1000 фаговых частиц. При инфицировании клетки-реципиента дефектной фаговой частицей ДНК клетки-донора «впрыскивается» в нее и рекомбинирует гомологичной рекомбинацией с гомологичным участком хромосомы-реципиента с образованием стабильного рекомбинанта. Этим типом трансдукции обладают Р-фаги;
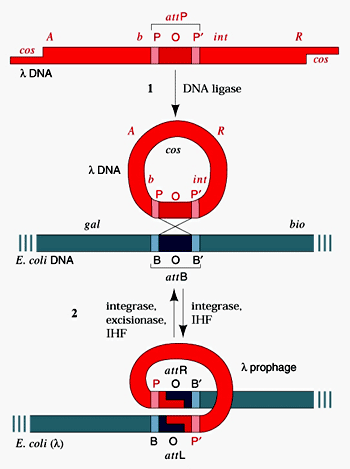
специфическая трансдукция – наблюдается в том случае, когда фаговая ДНК интегрирует в бактериальную хромосому с образованием профага. В процессе исключения
ДНК-фага из бактериальной хромосомы в результате случайного процесса захватывается прилегающий к месту включения фаговой ДНК фрагмент бактериальной хромосомы, становясь дефектным фагом. Так как большинство умеренных бактериофагов интегрирует в бактериальную хромосому в специфических участках, для таких бактериофагов характерен перенос в клетку-реципиент определенного участка бактериальной ДНК клетки-донора. ДНК дефектного фага рекомбинирует с ДНК клетки-реципиента сайт-специфической рекомбинацией. В частности, бактериофаг передает специфической трансдукцией gal-ген у Е. coli.
абортивнаятрансдукция – привнесенный фагом фрагмент ДНК бактерии-донора не включается в хромосому бактерии-реципиента, а располагается в её цитоплазме и может в таком виде функционировать. Во время деления бактериальной клетки трансдуцированный фрагмент ДНК-донора может передаваться только одной из двух дочерних клеток, т.е. наследоваться однолинейно и в конечном итоге утрачиваться в потомстве.
Трансдукция обнаружена у E.coli, B. subtilis, сальмонелл, холерного вибриона и др. Передаются самые различные свойства бактерий: устойчивость к антибиотикам, синтез факторов роста, сбраживание углеводов, синтез пенициллиназы и др.
⇐ Предыдущая1234
Дата публикования: 2014-12-10; Прочитано: 871 | Нарушение авторского права страницы
studopedia.org — Студопедия.Орг — 2014-2018 год.(0.003 с)…
определение
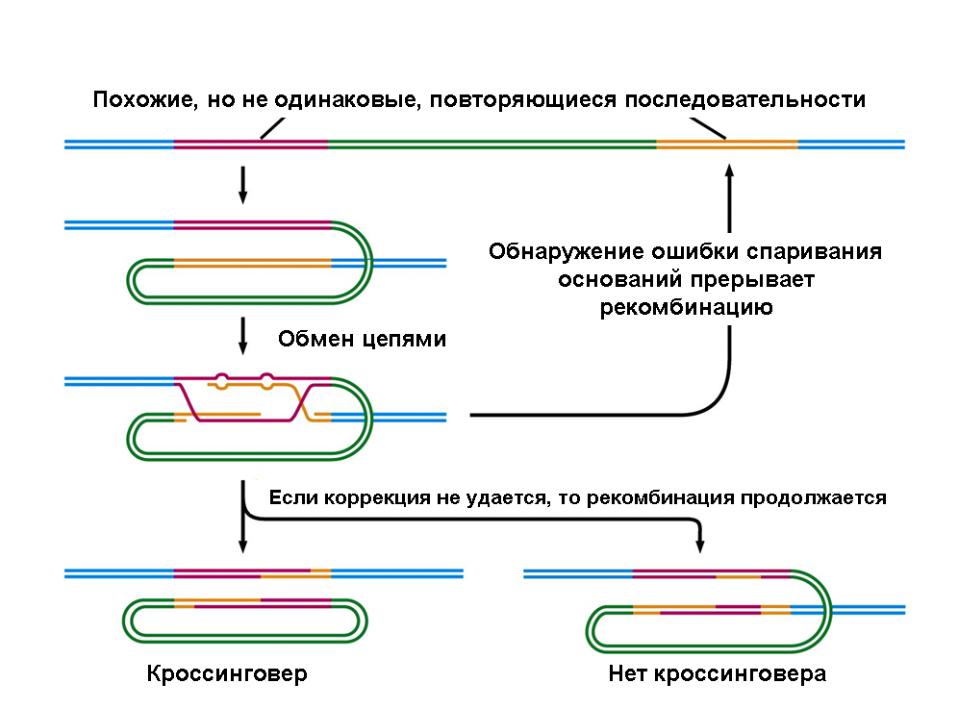
То, что мы называем гомологией в рекомбинации, не обязательно указывает на эволюционное происхождение участвующих молекул. Мы говорим больше о степени сходства в нуклеотидной последовательности.
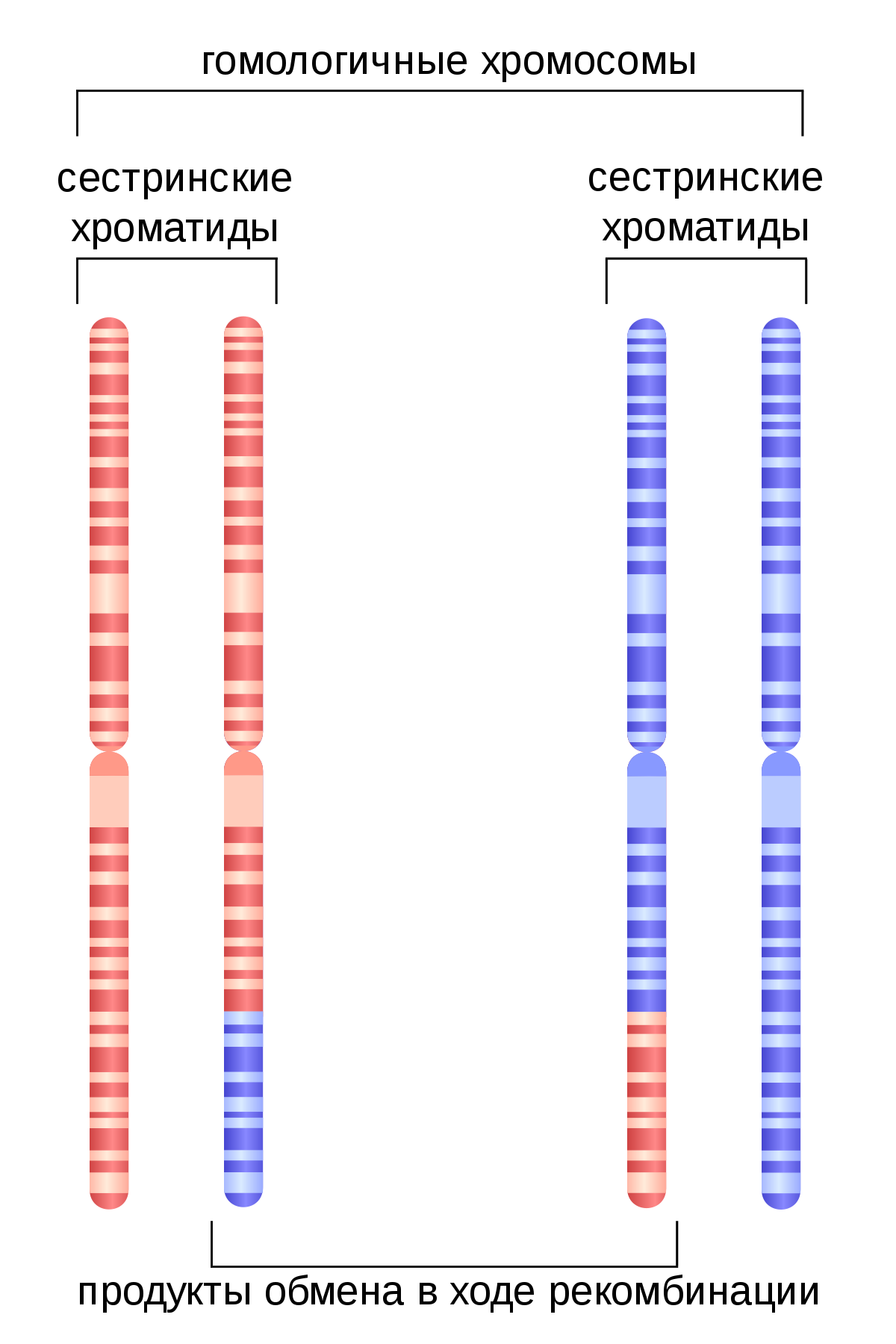
Например, нерепаративная рекомбинация происходит у эукариот во время процесса мейоза. Несомненно, не может быть большей гомологии, чем между парами хромосом в одной клетке.
Вот почему они называются гомологичными хромосомами. Однако существуют случаи, когда ДНК клетки обменивается материалом с чужеродной ДНК. Эти ДНК должны быть очень похожи на рекомбинацию, но они не обязательно должны иметь одного и того же предка (гомологию) для достижения этого..



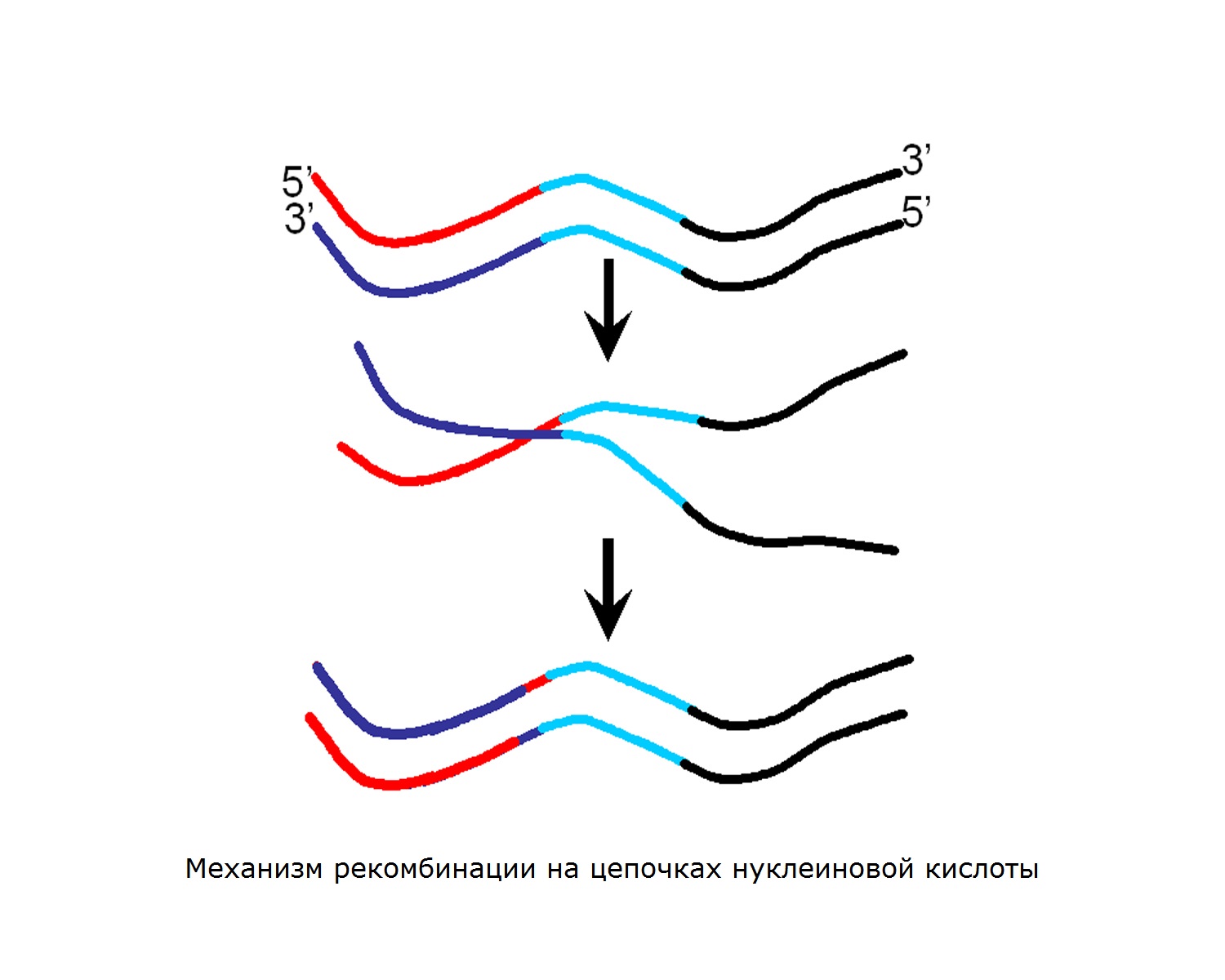
Хиазм и сшивание
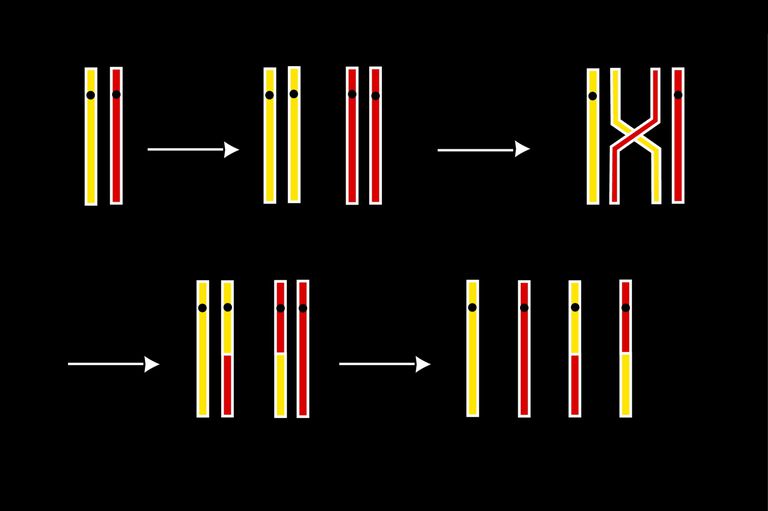
Сайт объединения и обмена между двумя молекулами ДНК называется хиазмом, а сам процесс сшивания. Во время скрещивания подтверждается обмен полос между участвующими ДНК..
Это создает коинтеграцию, которая представляет собой две молекулы ДНК, физически объединенные в одну. Когда коинтеграция «решена», генерируются две молекулы, обычно изменяемые (рекомбинантные).
«Решить» в контексте рекомбинации — разделить молекулы ДНК на компоненты коинтеграции..
Обзор

Электронная зонная структура полупроводникового материала.
Как и другие твердые тела, полупроводниковые материалы имеют электронную зонную структуру, определяемую кристаллическими свойствами материала. Распределение энергии между электронами описывается уровнем Ферми и температурой электронов. При абсолютном нуле температуры все электроны имеют энергию ниже уровня Ферми; но при ненулевых температурах уровни энергии заполняются согласно распределению Больцмана.
В нелегированных полупроводниках уровень Ферми находится в середине запрещенной зоны или запрещенной зоны между двумя разрешенными зонами, называемыми валентной зоной и зоной проводимости . Валентная зона, находящаяся непосредственно под запрещенной зоной, обычно почти полностью занята. Зона проводимости выше уровня Ферми обычно почти полностью пуста. Поскольку валентная зона почти заполнена, ее электроны неподвижны и не могут течь как электрический ток.
Однако, если электрон в валентной зоне приобретает достаточно энергии, чтобы достичь зоны проводимости (в результате взаимодействия с другими электронами , дырками , фотонами или самой колеблющейся кристаллической решеткой ), он может свободно течь среди почти пустой зоны проводимости. состояния. Кроме того, он также оставит отверстие, по которому может течь ток, точно как физическая заряженная частица.
Генерация носителей описывает процессы, посредством которых электроны набирают энергию и перемещаются из валентной зоны в зону проводимости, создавая два мобильных носителя; в то время как рекомбинация описывает процессы, посредством которых электрон в зоне проводимости теряет энергию и повторно занимает энергетическое состояние электронной дырки в валентной зоне.
Эти процессы должны сохранять как квантованную энергию, так и импульс кристалла , а колеблющаяся решетка играет большую роль в сохранении импульса, поскольку при столкновениях фотоны могут передавать очень небольшой импульс по сравнению с их энергией.
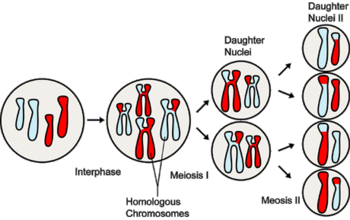
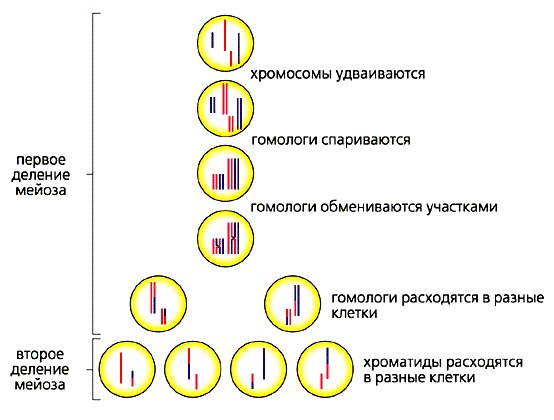
Мейоз
Хромосомный кроссовер включает обмен парными хромосомами, унаследованными от каждого из родителей. Мейоз, как основа рекомбинации генов, играет в этом процессе важную роль. Молекулярные модели этого процесса эволюционировали с годами по мере накопления соответствующих доказательств. Новая модель демонстрирует: два из четырех хроматидов, присутствующих в начале мейоза (профаза I), спарены друг с другом и способны взаимодействовать. В ней имеет место рекомбинация хромосом и генов. Однако объяснения адаптивной функции мейоза, которые фокусируются исключительно на пересечении, недостаточны для большинства событий обмена.

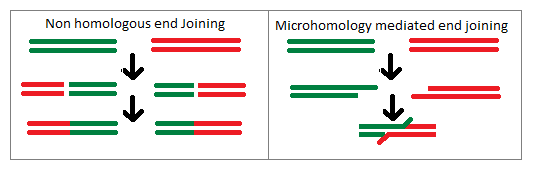
Рекомбинантное восстановление
Во время митоза и мейоза поврежденные различными экзогенными факторами ДНК могут быть спасены путем гомологичной стадии восстановления (ГСВ). У людей и грызунов дефицит генных продуктов, необходимых для ГСВ во время мейоза, вызывает бесплодие.
У бактерий трансформация — это процесс передачи генов, который обычно происходит между отдельными клетками одного и того же вида. Она предполагает интеграцию донорской ДНК в хромосому реципиента путем рекомбинации генов. Этот процесс является адаптацией для восстановления поврежденных клеток. Трансформация может принести пользу патогенным бактериям, позволяя восстанавливать повреждения ДНК, возникающие в воспалительной, окислительной среде, связанной с инфекцией хозяина.
Когда два или более вирусов, каждый из которых содержит смертельные геномные повреждения, заражают одну и ту же клетку-хозяина, геномы могут спариваться друг с другом и проходить ГСВ для получения жизнеспособного потомства. Этот процесс называется реактивацией множественности. Он изучался у нескольких патогенных вирусов.

Поколение перевозчика
Когда свет взаимодействует с материалом, он может либо поглощаться (генерируя пару свободных носителей или экситон ), либо стимулировать событие рекомбинации. Сгенерированный фотон имеет те же свойства, что и тот, который вызвал событие. Поглощение — это активный процесс в фотодиодах , солнечных элементах и других полупроводниковых фотодетекторах , тогда как вынужденное излучение является принципом работы в лазерных диодах .






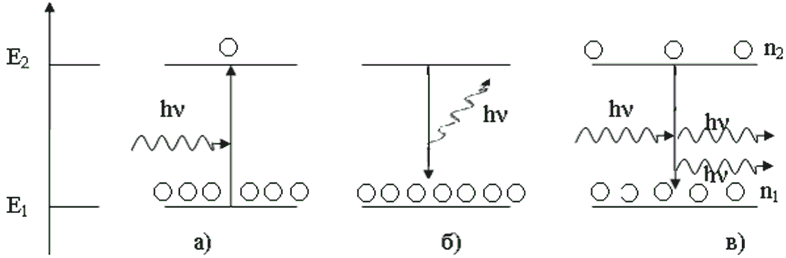


Помимо возбуждения светом, носители в полупроводниках могут также генерироваться внешним электрическим полем, например, в светодиодах и транзисторах .
Когда свет с достаточной энергией попадает на полупроводник, он может возбуждать электроны через запрещенную зону. Это генерирует дополнительные носители заряда, временно снижая электрическое сопротивление материалов. Эта более высокая проводимость в присутствии света известна как фотопроводимость . Это преобразование света в электричество широко используется в фотодиодах .
Функции и последствия гомологичной рекомбинации
Генетический материал постоянно подвергается повреждениям, вызванным эндогенными и экзогенными источниками, такими как, например, радиация..
Подсчитано, что клетки человека представляют значительное количество повреждений в ДНК, порядка от десятков до сотен в день. Эти повреждения должны быть исправлены, чтобы избежать потенциальных вредных мутаций, блокировок в репликации и транскрипции и повреждения на хромосомном уровне..
С медицинской точки зрения повреждение ДНК, которое не восстанавливается правильно, приводит к развитию опухолей и других патологий..
Гомологичная рекомбинация — это событие, которое позволяет репарации в ДНК, позволяя восстановить потерянные последовательности, используя в качестве матрицы другую цепь ДНК (гомологичную)..
Этот метаболический процесс присутствует во всех формах жизни, обеспечивая механизм высокой точности, который позволяет восстанавливать «разрывы» в ДНК, двухцепочечные разрывы и поперечные связи между цепями ДНК.
Одним из наиболее важных последствий рекомбинации является генерация новых генетических вариаций. Вместе с мутациями они являются двумя процессами, которые генерируют вариации в живых существах — помните, что вариации являются сырьем для эволюции.
Кроме того, он предоставляет механизм для перезапуска вилок репликации, которые были повреждены.
В бактериях
У бактерий нередки случаи горизонтального переноса генов. Они классифицируются как сопряжение, преобразование и преобразование. Здесь прокариоты берут ДНК из другого организма, и даже из разных видов.
Во время этих процессов происходит гомологичная рекомбинация между клеткой-реципиентом и клеткой-донором.
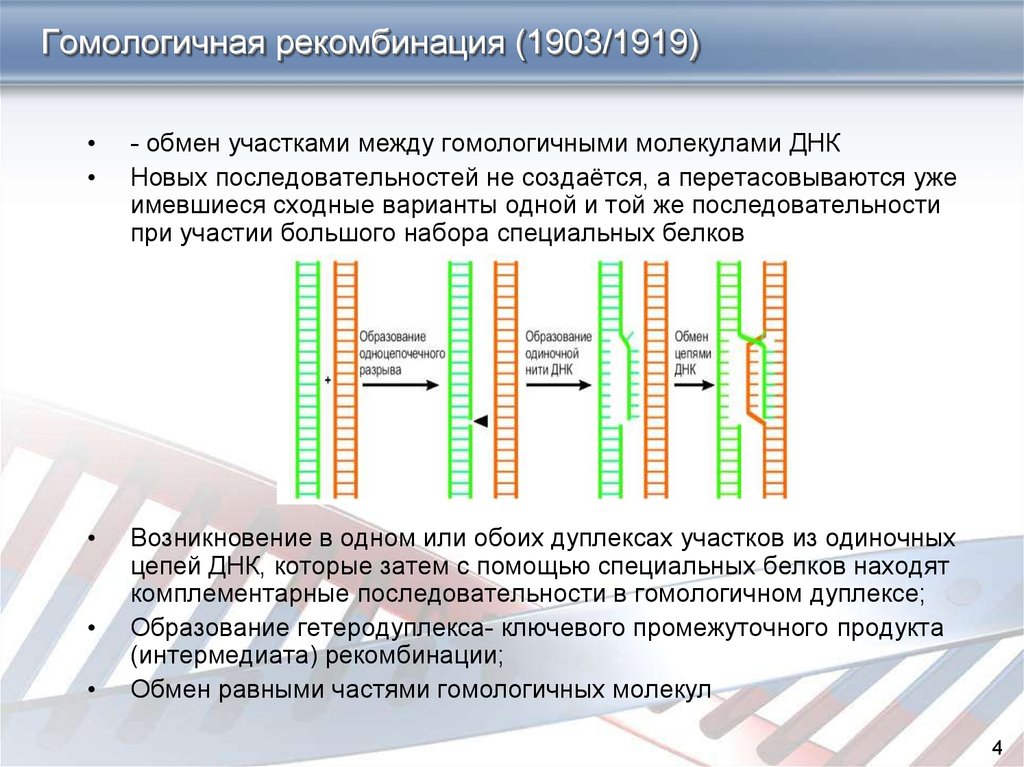
механизм
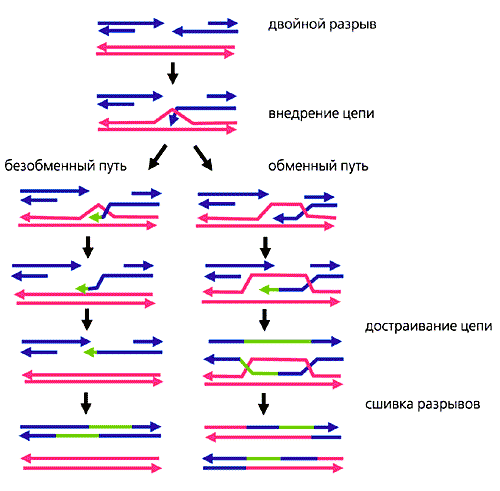
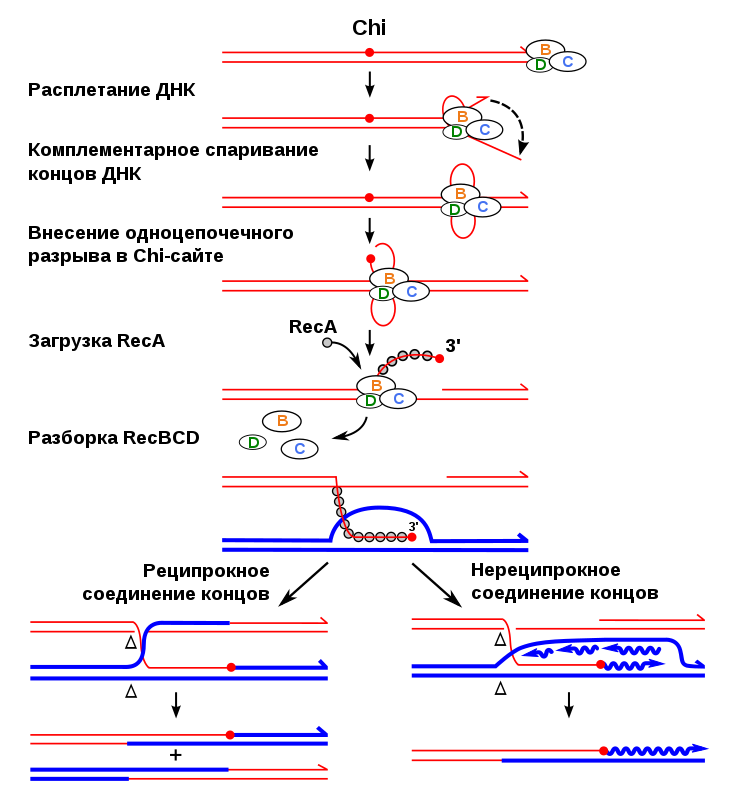
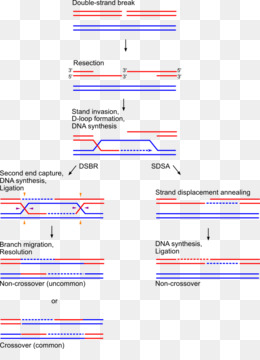
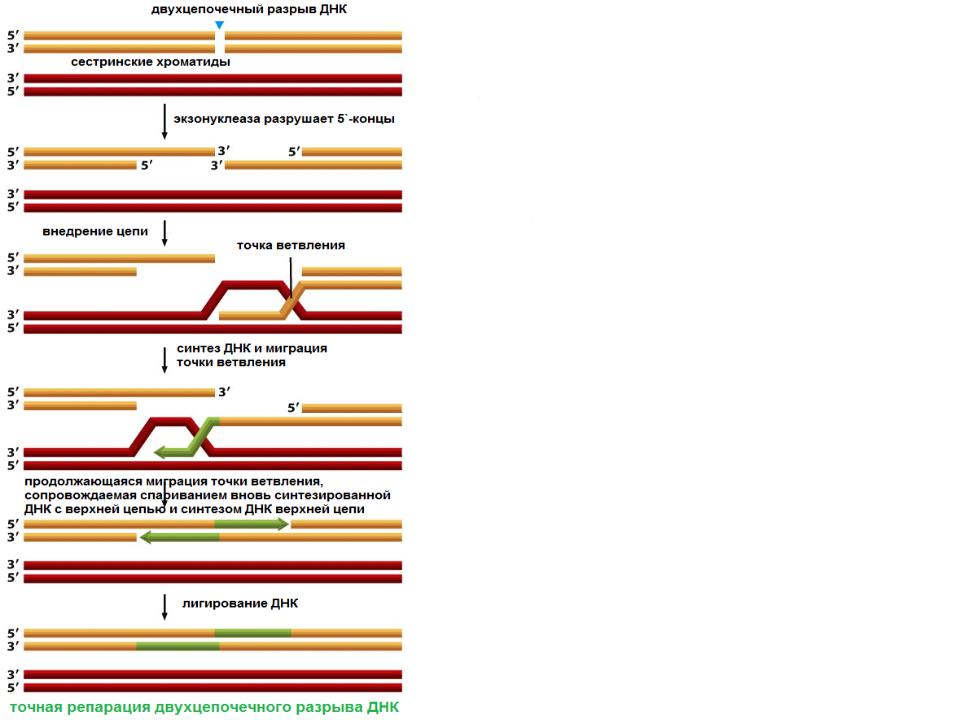
Гомологичная рекомбинация начинается с разрыва одной из цепей молекулы хромосомной ДНК. После этого происходит ряд этапов, катализируемых несколькими ферментами..
3′-конец, где происходит разрез, пронизан двойной гомологичной цепью ДНК. Процесс вторжения имеет решающее значение. С «гомологичной цепью» мы хотим ссылаться на части хромосом, которые имеют одинаковые гены в линейном порядке, хотя нуклеотидные последовательности не обязательно должны быть идентичными.
синапс
Эта инвазия нити размещает гомологичные хромосомы лицом друг к другу. Этот феномен столкновения нитей называется синапсисом (не путать с синапсами в нейронах, здесь этот термин используется с другим значением).
Синапс не обязательно подразумевает прямой контакт между обеими гомологичными последовательностями, ДНК может продолжать двигаться некоторое время, пока не найдет гомологичную часть. Этот процесс поиска называется гомологичным выравниванием.

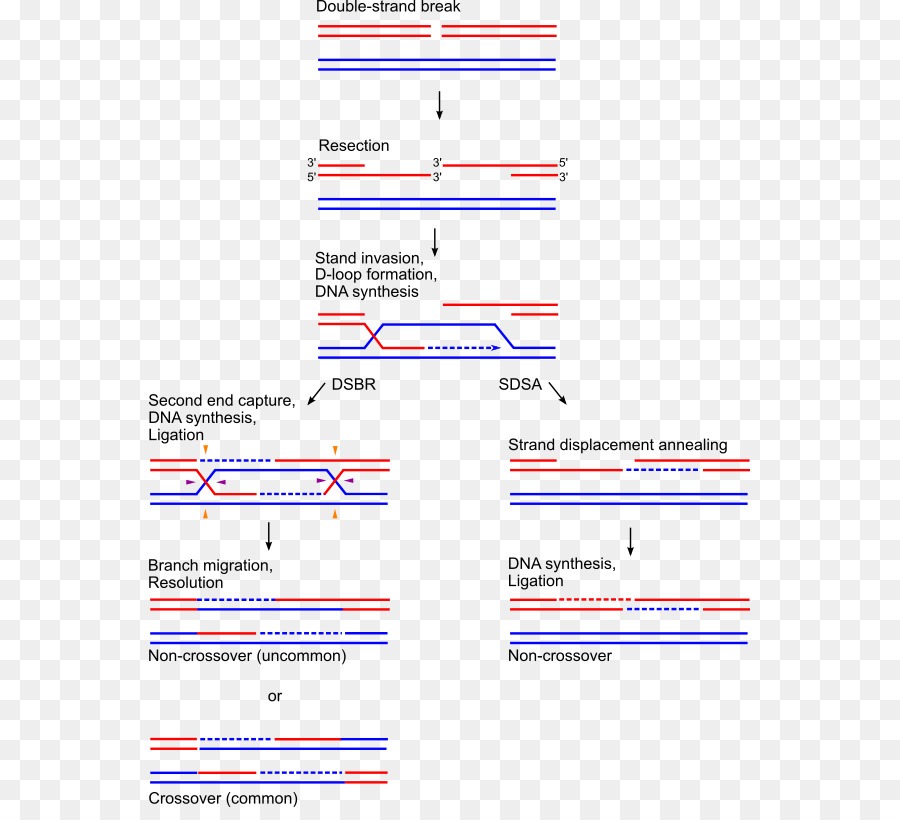


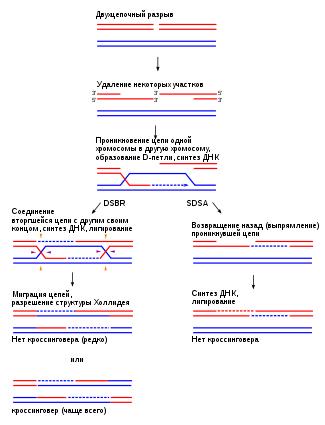
Формирование петли D
Затем происходит событие, называемое «вторжение в поток». Хромосома — это двойная спираль ДНК. При гомологичной рекомбинации две хромосомы ищут свои гомологичные последовательности. В одной из спиралей нити разделяются, и эта нить «проникает» в структуру двойной спирали, образуя структуру, называемую петлей D.
Цепочка D-петли была смещена вторжением нити, которая представляет разрыв и соединяется с комплементарной нитью исходной двойной спирали.
Формирование Холлидей Союзов
Следующим шагом является формирование Холлидейских перекрестков. Здесь концы обмениваемых потоков связаны между собой. Этот союз имеет возможность двигаться в любом направлении. Союз может быть разбит и образован несколько раз.
Последний процесс рекомбинации — разрешение этих соединений, и есть два пути или пути, которыми клетка достигает этого. Одним из них является расщепление соединения или процесс, называемый растворением, типичным для эукариотических организмов..
В первом механизме разрыв объединения Холлидей восстанавливает две цепи. В другом случае «роспуска» происходит своего рода коллапс в союзе.
Белки участвуют
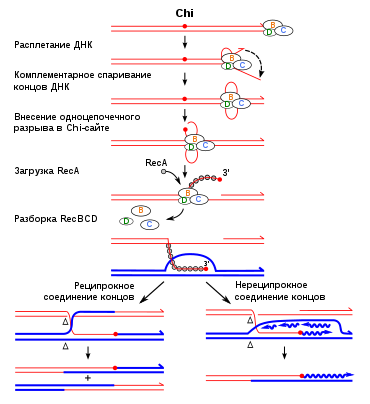
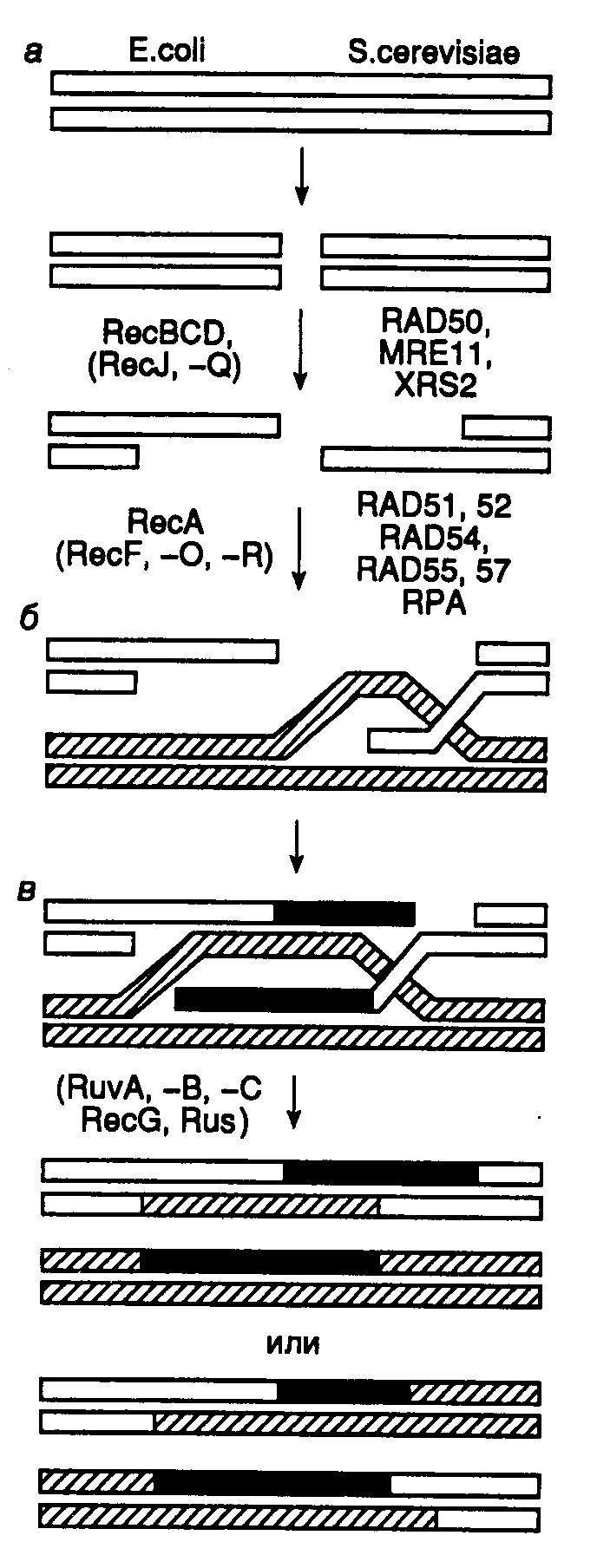
Важный белок процесса рекомбинации называется Rad51 в эукариотических клетках, а RecA в Кишечная палочка. Он работает на разных фазах рекомбинации: до, во время и после синапса.
Белок Rad51 облегчает формирование физической связи между вторгающейся ДНК и закаленной ДНК. В этом процессе генерируется гетеродуплексная ДНК.
Rad51 и его гомолог RecA катализируют поиск гомологичной ДНК и обмен цепей ДНК. Эти белки обладают способностью объединяться в одну полосу ДНК.
Существуют также паралогические гены (возникшие в результате дупликации генов в линии организмов) Rad51, называемые Rad55 и Rad57. У людей было идентифицировано пять генов паралога Rad51, названных Rad51B, Rad51C, Rad51D, Xrcc2 и Xrcc3..