Дедуктивный метод Копечки-Гейни
Метод Браун-Бланке в его классическом понимании основывается на характерных видах и однозначной дихотомии. Этот подход оправдал себя при классификации естественных богатых видами сообществ, когда выделялся ряд синтаксонов, с которыми соотносилась часть сообществ, а другие рассматривались как переходные. Но при классификации такого метода гиперконтинуальной рудеральной растительности возникли сложности, связанные с широкой экологической амплитудой, эврибионтностью входящих в неё видов. Это приводило к невозможности выделить характерные для той или иной ассоциации и даже союза виды. Потому в 1974 году чешские ботаники К. Копечки и С Гейни предложили так называемый дедуктивный метод классификации синантропной растительности, который заключается в том, что вместе с ассоциациями выделяются сообщества, которые подчиняются непосредственно классу или порядку, или одновременно двум высшим синтаксонам на основе представления в них диагностических видов высших единиц. Различают базальные сообщества (сформированные «своим» доминантом) и дериватные (доминант которых является диагностическим видом другого синтаксона).
Основные высшие синтаксоны растительности средней полосы европейской части России
-
Лесная растительность:
- класс Querco-Fagetea Br.-Bl. et Vlieg. 1937 — мезофитные и мезоксерофитные широколиственные листопадные леса на богатых почвах
- класс Vaccinio-Piceetea Br.-Bl. 1939 — бореальные сосновые леса на бедных кислых почвах с развитым моховым покровом
- класс Alnetea glutinosae Br.-Bl. et R.Tx. 1943 — низинные эвтрофные заболоченные леса с преобладанием ольхи чёрной и березы пушистой на торфяных и торфяно-минеральных почвах
- класс Salicetea purpureae Moor 1958 — пойменные прирусловые ивняково-тополёвые леса и кустарниковые заросли
-
Луговая растительность:
- класс Molinio-Arrhenatheretea R.Tx. 1937 em. R.Tx. 1970 — вторичные послелесные луга, формирующиеся на месте широколиственных лесов с довольно богатыми незасолёнными почвами
- класс Sedo-Scleranthethea Br.Bl. 1955 — травянистые сообщества на слаборазвитых песчаных почвах
-
Растительность, сформированная на нарушенных участках:
- класс Epilobietea angustifolii R.Tx. et Prsg. in R.Tx. 1950 — растительность вырубок и гарей
- класс Galio-Urticetea Passage 1967 — естественные и антропогенные нитрофильные сообщества затенённых мест, засорённых участков, опушек в лесопарках, в поймах рек и ручьёв
- класс Plataginetea majoris R.Tx. et Prsg. in R.Tx. 1950 — сообщества низкорослых, устойчивых к вытаптыванию и выпасу мезофитов и гигрофитов во дворах, вдоль дорог, на спортивных площадках
- класс Chenopodietea Br.-Bl. 1952 em. Lohm. J. et R.Tx. 1961 ex Matusz. 1962 — сообщества однолетников, представляющих собой начальные стадии восстановительных сукцессий после нарушений и сорнополевые сообщества
- класс Artemisietea vulgaris Lohm., Prsg. et Tx. in Tx. 1950 — рудеральные сообщества высокорослых двухлетних и многолетних видов
- класс Agropyretea repentis Oberd., Th.Muller et Gors in Oberd. et al. 1967 — рудеральные сообщества с преобладанием многолетних злаков, представляющих собой предыдущую перед лугами стадию восстановительных сукцессий
- класс Bidentetea tripartiti R.Tx., Lohm. et Prsg 1950 — синантропные сообщества с преобладанием однолетних видов нарушенных переувлажнённых мест
- класс Secalietea Br.-Bl. 1951 — сегетальные (сорнополевые) сообщества зерновых культур
- класс Robinietea Jurko ex Sofron 1980 — городская спонтанная растительность и сообщества искусственных древесных насаждений
-
Прибрежно-водная растительность:
- класс Phragmiti-Magnocaricetea Klika in Klika et Novak 1941 — водные и околоводные сообщества прикреплённых ко дну и возвышающихся над водой растений
- класс Potametea R.Tx. et Prsg — сообщества прикреплённых ко дну растений с плавающими на поверхности или погружёнными в толщу воды листьями
- класс Lemnetea minoris R. Tx. 1955 — сообщества свободно плавающих на поверхности и в толще воды незакреплённых растений
- класс Charetea (Fukarek 1961 n.n.) Krausch 1964 — сообщества харовых водорослей
- класс Utricularietea intermedio-minoris Den Hartog et Segal 1943 em. Pietsch 1965 — пузырчатко-моховые сообщества дистрофных вод, обычно в мочажинах и озёрах на верховых и переходных болотах
Определение
История
На вопрос, что называть растением, нет однозначного ответа. Первым на этот вопрос попытался ответить древнегреческий философ и учёный Аристотель, поместив растения в промежуточное состояние между неодушевлёнными предметами и животными. Он определил растения как живые организмы, которые не способны самостоятельно передвигаться (в противоположность животным). Позднее были открыты бактерии и археи, которые никак не подпадали под общепринятое понятие растений. Уже во второй половине XX века грибы и некоторые типы водорослей были выделены в отдельные категории, поскольку не имеют сосудистой и корневой системы, которая присутствует у других растений.
Определяющие признаки
- Наличие плотной, не пропускающей твёрдые частицы, клеточной оболочки (как правило, состоящей из целлюлозы)
- Растения — продуценты. Они производят органические вещества с помощью углекислого газа и энергии солнца в процессе фотосинтеза. Грибы и бо́льшая часть бактерий в последнее время относятся к отдельным царствам. Раньше грибы и бактерии считались растениями.
- Цианобактерии, или синезелёные водоросли, для которых, как и для большинства растений свойственен фотосинтез, согласно современным классификациям также не относятся к растениям (цианобактерии включены в домен Бактерии в ранге отдела).
- Другие признаки растений — неподвижность, постоянный рост, чередование поколений и другие — не являются уникальными, но в целом позволяют отличить растения от других групп организмов.
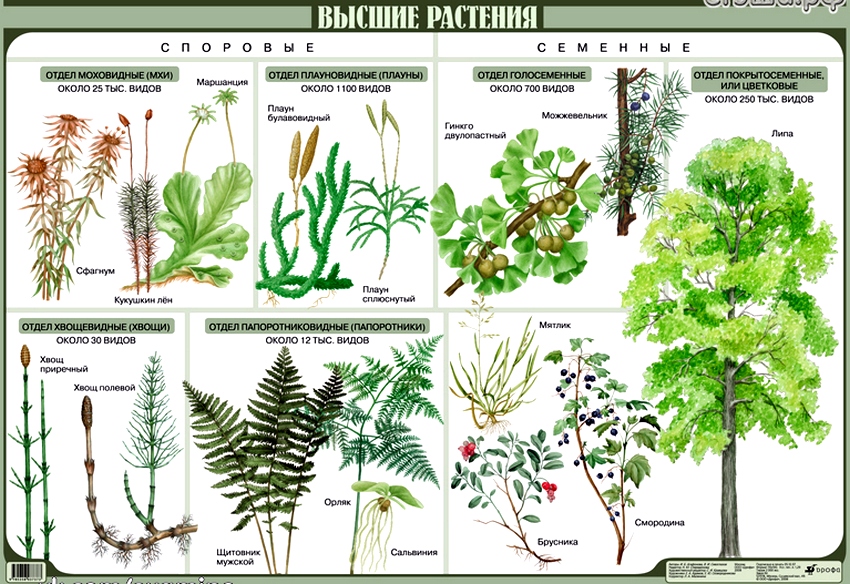
Разнообразие
По состоянию на начало 2010 года, по данным Международного союза охраны природы (IUCN), было описано около 320 тысяч видов растений, из них около 280 тысяч видов цветковых, 1 тысяча видов голосеменных, около 16 тысяч мохообразных, около 12 тысяч видов высших споровых растений (Плауновидные и Папоротникообразные). Однако, это число увеличивается, так как постоянно открываются новые виды.
| Отделы | Русскоеназвание | Числовидов | |
|---|---|---|---|
| Водоросли | Chlorophyta | Зелёные водоросли | 13 000 — 20 000 |
| Charophyta | Харофиты | 4000—6000 | |
| Мохообразные | Marchantiophyta | Печёночные мхи | 6000—8000 |
| Anthocerotophyta | Антоцеротовые мхи | 100—200 | |
| Bryophyta | Моховидные | 10 000 | |
| Сосудистые споровые | Lycopodiophyta | Плауновидные | 1200 |
| Polypodiophyta | Папоротниковидные | 11 000 | |
| Семенные растения | Cycadophyta | Саговниковидные | 160 |
| Ginkgophyta | Гинкговидные | 1 | |
| Pinophyta | Хвойные | 630 | |
| Gnetophyta | Гнетовидные | 70 | |
| Magnoliophyta | Цветковые растения | 281 821 |
Значение слова Растительность по словарю Брокгауза и Ефрона:
Растительность — Слову Р. Визнер предает специальный смысл, противополагая это слово термину флора. Приспособление к климату и почве накладывает особый отпечаток на характер растительного покрова, давая тот или иной тип Р., независимо от систематического положения составляющих данную Р. видов. Так, молочаи жаркой области пустынь по своему биологическому характеру чрезвычайно сходны с кактусами, хотя в систематическом отношении не имеют ничего общего с ними. Таким образом, облик Р. пустынь будет одинаков, составят ли его кактусы или кактусовидные молочаи. Наоборот, флора в этих обо их случаях будет резко различна, так как под флорой понимают не данный биологический тип растительного покрова, а его определенный систематический состав. Таким образом, характеристика Р. сводится к установлению биологических признаков растительного покрова, характеристика же флоры определяется точным установлением составляющих ее растительных видов. Вторым ярким примером одинаковости Р. при резком различии флоры могут явиться Новая Земля и аналогично расположенные Малуиновы острова в Атлантическом океане, обладающие приблизительно таким же климатом, как и Новая Земля. Характер флоры в обоих этих случаях резко различен: из 193 видов новоземельской флоры 10 приходятся на род Saxifraga, 20 относятся к семейству крестоцветных, среди них найдено 10 видов Draba, найдена Dryas octopetala и пр. Напротив, на Малуиновых островах флора насчитывает лишь 135 видов. среди них совсем нет Saxifraga и Dryas. семейство крестоцветных имеет здесь лишь трех представителей зато на этих островах есть ряд растений, чуждых Новой Земле. Таким образом, флора здесь и там совершенно различна. напротив, характер Р. один и тот же. Те же тундры, то же отсутствие деревьев, то же преобладание травянистых многолетников с коротким вегетационным периодом характеризует Р. обеих местностей.
Смысл понятия «растительность»
Понятие «растительность» (растительный покров) означает исторически сформированное объединение растительных сообществ (фитоценозов) на определённой территории.
Растительный покров представляет собой совокупность фитоценозов сообществ растений, которые населяют планету и её регионы. Основной единицей классификации растительности есть растительная ассоциация. Классификация растительности базируется на экологических, флористических, генетических, фитоценологических принципах.
Растительность — это не случайный набор видов, а закономерное их сочетание, которое возникло под влиянием экологических факторов на протяжении длительного времени. Растительность является чрезвычайно важным компонентом биосферы, поскольку она связана с особенностями климата, водного режима, структурой почв, рельефом, с животным миром. Распределение растительности в большинстве случаев имеет зональный характер и связано с климатическими поясами (распределением энергии Солнца). В горах распределение растительности подчинено высотной зональности.
Пример 4
Высокогорная растительность Кавказа, растительность умеренного пояса, растительность заливных лугов.
Основные критерии характеристики растительности:
видовой состав;
количество экземпляров на определённой территории.
Существуют 4 типа растительности:
- древесный,
- травянистый,
- пустынный,
- планктонный.
В зависимости от места произрастания на нашей планете выделяют растительность лесов, гилеи, саванны, мангры, высокогорную растительность, сухие редколесья, кустарники, злаковники ( степи и луга), полупустыни, пустыни, тундры, умеренных и высоких широт, вересковники, низинные болота, верховые болота, растительность морей.
Появление и эволюция
Архейская эра (2500—3800 млн лет назад)
Судя по палеонтологическим находкам, разделение живых существ на царства произошло более 3 млрд лет назад. Первыми автотрофными организмами стали фотосинтезирующие бактерии (сейчас они представлены пурпурными и зелёными бактериями, цианобактериями). В частности, в мезоархее (2800—3200 млн лет назад) уже существовали цианобактериальные маты.










Протерозойская эра (570—2500 млн лет назад)
Единой, отвечающей на все вопросы, теории происхождения эукариотических фотоавтотрофных организмов (растений) пока нет. Одна из них (теория симбиогенеза) предполагает возникновение эукариотических фототрофов как переход эукариотической гетеротрофной амёбовидной клетки к фототрофному типу питания через симбиоз с фотосинтезирующей бактерией, которая впоследствии превратилась в хлоропласт. Согласно этой теории, таким же образом возникают и митохондрии из аэробных бактерий. Так появляются водоросли — первые настоящие растения. В протерозойскую эру широко развиваются одноклеточные и колониальные синезелёные водоросли, появляются красные и зелёные водоросли.
Палеозойская эра (230—570 млн лет назад)
В конце Силура (405—440 млн лет назад) на Земле происходят интенсивные горообразовательные процессы, приведшие к возникновению Скандинавских гор, гор Тянь-Шань, Саян, а также к обмелению и исчезновению многих морей. В результате некоторые водоросли (сходные с современными харовыми водорослями) выходят на сушу и заселяют литорали и супралиторали, что стало возможным благодаря деятельности бактерий и цианобактерий, образовавших на поверхности суши почвенный субстрат. Так возникают первые высшие растения — риниофиты. Особенность риниофитов заключается в появлении тканей и их дифференцировки на покровные, механические, проводящие и фотосинтезирующие. Это было спровоцировано резким отличием воздушной среды от водной. В частности:
- повышенной солнечной радиацией, для защиты от которой у первых наземных растений должен был выделяться и откладываться на поверхности кутин, что и было первым этапом формирования покровных тканей (эпидермы);
- откладывание кутина делает невозможным поглощение влаги всей площадью (как у водорослей), что приводит к изменению функции ризоидов, которые теперь не только прикрепляют организм к субстрату, но и поглощают из него воду;
- разделение на подземную и надземную части спровоцировало необходимость доставки минеральных веществ, воды и продуктов фотосинтеза по всему организму, реализованную появившимися проводящими тканями — ксилемой и флоэмой;
- отсутствие выталкивающей силы воды и соответственно невозможность плавать, в ходе конкуренции видов за солнечный свет, привело к появлению механических тканей с целью «приподняться» над соседями, ещё одним фактором было улучшенное освещение, активизировавшее процесс фотосинтеза и приведшее к избытку углерода, что и позволило образоваться механическим тканям;
- в ходе всех вышеперечисленных ароморфозов фотосинтезирующие клетки выделяются в отдельную ткань.
Древнейшее известное наземное растение — куксония. Куксония обнаружена в 1937 г. в силурийских песчаниках Шотландии (возраст порядка 415 млн лет). Дальнейшая эволюция высших растений разделилась на две линии: гаметофитную (моховидные) и спорофитную (сосудистые растения). Первые голосеменные растения появляются в начале Мезозоя (примерно 220 млн лет назад). Первые покрытосеменные (цветковые) возникают в юрском периоде.
Строение растений
Некоторые растения имеют очень сложное строение, но некоторые представлены одноклеточными организмами. Например: хлорелла, хламидомонада и т. д.
Для клеток растений характерен большой относительный размер (иногда до нескольких сантиметров), наличие жёсткой клеточной оболочки из целлюлозы, присутствие хлоропластов и крупной центральной вакуоли, позволяющей регулировать тургор. Во время деления перегородка образуется за счёт слияния многочисленных пузырьков (фрагмопласт). Сперматозоиды растений дву- (у мохообразных и плауновидных) или многожгутиковые (у остальных папоротникообразных, саговниковых и гинкговых), причём ультраструктура жгутикового аппарата очень похожа на таковую в жгутиковых клетках харовых водорослей (отдел Зелёные водоросли).
Клетки растений объединяются в ткани. Ткани растений характеризуются отсутствием межклеточного вещества, большим количеством мёртвых клеток (некоторые ткани, такие как склеренхима и пробка, состоят в основном из мёртвых клеток), а также тем, что, в отличие от животных, ткань растения может состоять из разных типов клеток (например, ксилема состоит из водопроводящих элементов, волокон древесины и древесинной паренхимы).
Большинство растений характеризуется значительным расчленением тела. Существуют несколько типов организации тела растений: талломный, при котором отдельные органы не выделяются и тело представляет собой зелёную пластину (некоторые мохообразные, заростки папоротников), листостебельный, при котором тело представляет собой побег с листьями (корни отсутствуют; большинство мохообразных), и корнепобеговый, когда тело делится на корневую и побеговую систему. Побег большинства растений состоит из осевой части (стебля) и боковых фотосинтезирующих органов (листьев), которые могут возникать либо как выросты внешних тканей стебля (у мохообразных), либо как следствие слияния укороченных боковых ветвей (у папоротникообразных). Зачаток побега принято считать особым органом — почкой.
Клетки растений содержат механорецепторы, корневые системы высших растений обмениваются органическими и неорганическими веществами с почвенными грибами.
Литература
- Александрова В. Д. Классификация растительности. Обзор принципов классификации и классификационных систем в различных геоботанических школах. — Л.: Наука, 1969. — 275 с.
- Василевич В. И. Что считать естественной классификацией // Философские проблемы современной биологии. — М.-Л.: Наука, 1966. — С. 177—190.
- Ипатов В. С., Кирикова Л. А. Фитоценология. — СПб.: Изд-во СПбГУ, 1998. — 314 с.
- Ишбирдин А. Р., Абрамова Л. М. Опыт классификации синантропной флоры на основе системы Браун-Бланке // Бюлл. МОИП. Отд. биол. 1990. Т. 95. Вып. 3. — С. 69—75.
- Методы выделения растительных ассоциаций. — Л.: Наука, 1971.
- Миркин Б. М., Наумова Л. Г., Соломещ А. И. Современная наука о растительности. — М.: Логос, 2001. — 264 с.
- Миркин Б. М., Розенберг Г. С., Наумова Л. Г. Словарь понятий и терминов современной фитоценологии. — М.: Наука, 1989. — 223 с.
- Миркин Б. М. Закономерности формирования растительности речных пойм. — М.: Наука, 1974.
- Миркин Б. М. Теоретические основы современной фитоценологии. — М.: Наука, 1985.
- Норин Б. Н. Растительный покров: ценотическая организация и объекты классификации // Ботанический журнал, 1983, т. 68, № 11. — С. 1449—1455.
- Погребняк П. С. и др. Основы лесной типологии. — Киев, 1944, 1955.
- Braun-Blanquet J. Pflanzensociologie. — Wien — New York, 1964. — 865 p.
- Kopecky K., Hejny S. A new approach to the classification of anthropogenic plant communities // Vegetatio. — 1974. — V. 29. — P. 17—20.
- Matuszkiewicz W. Przewodnik do oznaczania zbiorowisk roślinnych Polski. — Warszawa: PWN, 2001. — 321 s.
Флора и растительность как предмет изучения
Понятия «флора» и «растительность» необходимо чётко различать. В отличии от флоры, растительность характеризуется не видовым составом растений, а численностью и сочетанием видов, их пространственной структурой и динамикой.









Пример 5
Флора умеренной зоны Северного полушария богато представлена видами растений из семейств Вербовые, Осоковые, Злаковые, Лютиковые, Астровые, Сосновые, Бобовые, Капустные, Розовые и др., а растительность — лесными, степными, пустынными, луговыми, болотными, тундровыми и другими сообществами.
Растительность — это предмет изучения науки геоботаники, или фитоценологии. Флора же есть объектом исследования географии растений. Между фитоценологией и географией растений невозможно провести чёткую границу. Эти науки взаимопроникающие и дополняющие друг друга. В то же время они, неразрывно связаны с экологией.
Разнообразие
По состоянию на начало 2010 года, по данным Международного союза охраны природы (IUCN), было описано около 320 тысяч видов растений, из них около 280 тысяч видов цветковых, 1 тысяча видов голосеменных, около 16 тысяч мохообразных, около 12 тысяч видов высших споровых растений (Плауновидные и Папоротникообразные). Однако, это число увеличивается, так как постоянно открываются новые виды.
| Отделы | Русскоеназвание | Числовидов | |
|---|---|---|---|
| Водоросли | Chlorophyta | Зелёные водоросли | 13 000 — 20 000 |
| Charophyta | Харофиты | 4000—6000 | |
| Мохообразные | Marchantiophyta | Печёночные мхи | 6000—8000 |
| Anthocerotophyta | Антоцеротовые мхи | 100—200 | |
| Bryophyta | Моховидные | 10 000 | |
| Сосудистые споровые | Lycopodiophyta | Плауновидные | 1200 |
| Polypodiophyta | Папоротниковидные | 11 000 | |
| Семенные растения | Cycadophyta | Саговниковидные | 160 |
| Ginkgophyta | Гинкговидные | 1 | |
| Pinophyta | Хвойные | 630 | |
| Gnetophyta | Гнетовидные | 70 | |
| Magnoliophyta | Цветковые растения | 281 821 |
Растительность в Энциклопедическом словаре:
Растительность — совокупность растительных сообществ (фитоценозов),населяющих какую-либо территорию. Распределение растительностиопределяется в основном общеклиматическими условиями и подчиняется законамзональности и поясности. Основные классификационные единицы — типрастительности, формация, ассоциация. От растительности отличают флору -совокупность систематических единиц (видов, родов, семейств) на даннойтерритории. В результате деятельности человека естественные сообществарастений в значительной степени заменены антропогенной растительностью.
Примечания
- ↑ Шипунов А. Б. Растения // Биология: Школьная энциклопедия / Белякова Г. и др. — М.: БРЭ, 2004. — 990 с. — ISBN 5-85270-213-7.
- ↑
- Van den Hoek, C., D. G. Mann, & H. M. Jahns, 1995. Algae:An Introduction to Phycology. pages 343, 350, 392, 413, 425, 439, & 448 (Cambridge: Cambridge University Press). ISBN 0-521-30419-9
- Van den Hoek, C., D. G. Mann, & H. M. Jahns, 1995. Algae:An Introduction to Phycology. pages 457, 463, & 476. (Cambridge: Cambridge University Press). ISBN 0-521-30419-9
- Crandall-Stotler, Barbara. & Stotler, Raymond E., 2000. «Morphology and classification of the Marchantiophyta». page 21 in A. Jonathan Shaw & Bernard Goffinet (Eds.), Bryophyte Biology. (Cambridge: Cambridge University Press). ISBN 0-521-66097-1
- Schuster, Rudolf M., The Hepaticae and Anthocerotae of North America, volume VI, pages 712—713. (Chicago: Field Museum of Natural History, 1992). ISBN 0-914868-21-7.
- Buck, William R. & Bernard Goffinet, 2000. «Morphology and classification of mosses», page 71 in A. Jonathan Shaw & Bernard Goffinet (Eds.), Bryophyte Biology. (Cambridge: Cambridge University Press). ISBN 0-521-66097-1
- ↑ Raven, Peter H., Ray F. Evert, & Susan E. Eichhorn, 2005. Biology of Plants, 7th edition. (New York: W. H. Freeman and Company). ISBN 0-7167-1007-2.
- Gifford, Ernest M. & Adriance S. Foster, 1988. Morphology and Evolution of Vascular Plants, 3rd edition, page 358. (New York: W. H. Freeman and Company). ISBN 0-7167-1946-0.
- Taylor, Thomas N. & Edith L. Taylor, 1993. The Biology and Evolution of Fossil Plants, page 636. (New Jersey: Prentice-Hall). ISBN 0-13-651589-4.
Жизненный цикл растений
Размножение
Для растений характерны два вида размножения: половое и бесполое. Для высших сосудистых растений единственной формой полового процесса является оогамия. Из форм бесполого размножении широко распространено вегетативное размножение.
Кроме вегетативных, растения имеют специализированные генеративные органы, строение которых связано с протеканием жизненного цикла. В жизненном цикле растений чередуется половое, гаплоидное поколение (гаметофит) и бесполое, диплоидное поколение (спорофит). На гаметофите образуются половые органы — мужские антеридии и женские архегонии (отсутствуют у некоторых гнетовых и у покрытосеменных). Сперматозоиды (их нет у хвойных, гнетовых и покрытосеменных) оплодотворяют находящуюся в архегонии яйцеклетку, в результате образуется диплоидная зигота. Зигота формирует зародыш, который постепенно развивается в спорофит. На спорофите развиваются спорангии (часто на специализированных спороносных листьях, или спорофиллах). В спорангиях происходит мейоз, и образуются гаплоидные споры. У разноспоровых растений эти споры двух типов: мужские (из них развиваются гаметофиты только с антеридиями) и женские (из них развиваются гаметофиты, несущие только архегонии); у равноспоровых споры одинаковые. Из споры развивается гаметофит, и всё начинается сначала. Такой жизненный цикл имеют Мохообразные и Папоротникообразные, причём у первой группы в жизненном цикле доминирует гаметофит, а у второй — спорофит. У семенных растений картина усложняется за счёт того, что женский (несущий архегонии) гаметофит развивается прямо на материнском спорофите, а мужской гаметофит (пыльцевое зерно) должен быть доставлен туда в процессе опыления. Спорофиллы у семенных растений часто сложно устроены и объединяются в так называемые стробилы, а у покрытосеменных растений — в цветки, которые могут, в свою очередь, объединяться в соцветия. Кроме того, у семенных растений возникает специализированная, состоящая из нескольких генотипов структура — семя, которое можно условно отнести к генеративным органам. У покрытосеменных растений цветок после опыления созревает и формирует плод.
Примечания
- ↑ Шипунов А. Б. Растения // Биология: Школьная энциклопедия / Белякова Г. и др. — М.: БРЭ, 2004. — 990 с. — ISBN 5-85270-213-7.
- ↑
- Van den Hoek, C., D. G. Mann, & H. M. Jahns, 1995. Algae:An Introduction to Phycology. pages 343, 350, 392, 413, 425, 439, & 448 (Cambridge: Cambridge University Press). ISBN 0-521-30419-9
- Van den Hoek, C., D. G. Mann, & H. M. Jahns, 1995. Algae:An Introduction to Phycology. pages 457, 463, & 476. (Cambridge: Cambridge University Press). ISBN 0-521-30419-9
- Crandall-Stotler, Barbara. & Stotler, Raymond E., 2000. «Morphology and classification of the Marchantiophyta». page 21 in A. Jonathan Shaw & Bernard Goffinet (Eds.), Bryophyte Biology. (Cambridge: Cambridge University Press). ISBN 0-521-66097-1
- Schuster, Rudolf M., The Hepaticae and Anthocerotae of North America, volume VI, pages 712—713. (Chicago: Field Museum of Natural History, 1992). ISBN 0-914868-21-7.
- Buck, William R. & Bernard Goffinet, 2000. «Morphology and classification of mosses», page 71 in A. Jonathan Shaw & Bernard Goffinet (Eds.), Bryophyte Biology. (Cambridge: Cambridge University Press). ISBN 0-521-66097-1
- ↑ Raven, Peter H., Ray F. Evert, & Susan E. Eichhorn, 2005. Biology of Plants, 7th edition. (New York: W. H. Freeman and Company). ISBN 0-7167-1007-2.
- Gifford, Ernest M. & Adriance S. Foster, 1988. Morphology and Evolution of Vascular Plants, 3rd edition, page 358. (New York: W. H. Freeman and Company). ISBN 0-7167-1946-0.
- Taylor, Thomas N. & Edith L. Taylor, 1993. The Biology and Evolution of Fossil Plants, page 636. (New Jersey: Prentice-Hall). ISBN 0-13-651589-4.
Общие сведения о классификации растительности
Классификация растительности является центральной частью фитоценологии. Уровень её развития отображает уровень развития всей науки.










Синтаксономия применяет опыт таксономии, которая уже к началу XX столетия была достаточно развитой наукой. Сложность классификации растительности объясняется тем, что растительные сообщества в отличие от видов — это условности, объединённые в многомерный континуум. Кроме того, растительные сообщества обладают невысоким уровнем целостности, что ведёт к изменчивости архетипов (наборов диагностических признаков) синтаксонов, иногда очень значительной. Также в синтаксонах более низких рангов весьма редко представлены полностью диагностические виды высших единиц. Все это обусловило значительную сложность выделения синтаксономических единиц и установление диагнозов сообществ.
Основные подходы к принципам и методологии классификации
До начала 1970-х годов существовало множество подходов к классификации растительности, которые постепенно трансформировались в два основных подхода:
- доминантный (физиономический).
- эколого-флористический.
При доминантном подходе синтаксоны выделяются по доминантам отдельных ярусов растительных сообществ. Основными единицами доминантной классификации являются формация — совокупность сообществ с одним доминантом (например, формация дубовых лесов) и ассоциация, которая выделяется на основании доминантов различных ярусов (например, дубрава рябиново-кисличная). Этот подход является простым и целиком применимым к лесной растительности бореальных, суббореальных и субтропических лесов, то есть там, где в составе сообщества имеются несколько явных доминантов с сильными эдификаторными свойствами и где растительный покров обладает высокой степенью дискретности.
Но этот подход является неприменимым при классификации лугов, рудеральной растительности, растительности вырубок и гарей, где доминанты могут быстро меняться в разные годы и даже сезоны и обладают слабыми эдификаторными свойствами, что ведет к большой вариабельности видового состава сообществ, и растительный покров характеризуется континуальностью и гиперконтинуальностью. Потому все большее распространение приобретает флористический подход.
Возникновение эколого-флористического подхода к классификации связано с именем швейцарского геоботаника Ж. Браун-Бланке. Метод классификации растительности, названный его именем, основывается на группировке сообществ в соответствии с подобием флористического состава, который отображает экологические условия и стадию сукцессии, и установлении синтаксонов от более низких к высшим. Синтаксоны в системе Браун-Бланке выделяются на основании диагностических видов, среди которых отличают характерные, дифференцирующие и константные:
- Характерные виды встречаются только в одном синтаксоне или встречаются в этом синтаксоне чаще, чем в других;
- Дифференцирующие виды диагностируют границей своего ареала и входят в состав нескольких синтаксонов;
- Константные виды встречаются с высоким постоянством, но проходят через этот синтаксон «транзитом».
Виды, константные для более низких единиц, могут быть дифференцирующими или характерными для высших. Но есть мнение, что разница между дифференцирующими и характерными видами незначительная, и часто их объединяют в единую группу диагностических видов (в этом случае константные виды рассматриваются отдельно).
Создание названий синтаксонов, их видоизменение и отмена регулируется «Кодексом фитосоциологической номенклатуры», впервые опубликованным в 1976 году. Существуют следующие основные ранги синтаксономической классификации (сверху вниз): класс (окончание -etea), порядок (-etalia), союз (-ion), ассоциация (-etum). При выделении синтаксона указывают фамилию его автора и год утверждения.
Перечень всех синтаксонов, встречающихся на определенной территории, называется продромусом. Пример:
- класс: Vaccinio-Piceetea Br.-Bl. 1939. Д. В. (диагностические виды) Dicranum scoparium, Pleurozium schreberi, Hylocomium splendens, Melampyrum pratense, Ptilium crista-castrensis, Pyrola minor, Pyrola rotundifolia, Peltigera aphthosa, Trientalis europaea, Vaccinium myrtillus, Vaccinium uliginosum, Vaccinium vitis-idaea
- порядок: Cladonio-Vaccinietalia Kiell.-Lund 1967. Д. В. Pinus sylvestris, Cladonia rangiferina, Cladonia arbuscula, Dicranum fuscescens, Dicranum polysetum, Arctostaphylos uva-ursi, Empetrum nigrum ssp. nigrum, Ledum palustre
- союз: Dicrano-Pinion Libb. 1933. Д. В. Chimaphila umbellata, Diphasiastrum complanatum, Monotropa hypopitys, Pyrola chlorantha, Viscum album ssp. аustriacum
ассоциация: Peucedano-Pinetum W.Mat. (1962) 1973. Д. В. Chimaphila umbellate, Peucedanum oreoselinum, Convallaria majalis, Polygonatum odoratum, Pulsatilla patens, Scorzonera humilis, Solidago virgaurea
-
- союз: Dicrano-Pinion Libb. 1933. Д. В. Chimaphila umbellata, Diphasiastrum complanatum, Monotropa hypopitys, Pyrola chlorantha, Viscum album ssp. аustriacum
- порядок: Cladonio-Vaccinietalia Kiell.-Lund 1967. Д. В. Pinus sylvestris, Cladonia rangiferina, Cladonia arbuscula, Dicranum fuscescens, Dicranum polysetum, Arctostaphylos uva-ursi, Empetrum nigrum ssp. nigrum, Ledum palustre