Эпистаз. Примеры
Ген называют эпистатическим (от греч. еpi — над), если его присутствие подавляет эффект какого-либо гена, находящегося в другом локусе. Эпистатические гены иногда называют ингибирующими генами, а те гены, действие которых ими подавляется, — гипостатическими.
Гены, подавляющие действие других генов, называются супрессорами или ингибиторами.
Они могут быть как доминантными, так и рецессивными. Гены-супрессоры известны у животных, растений и микроорганизмов. Обычно они обозначаются I или S.
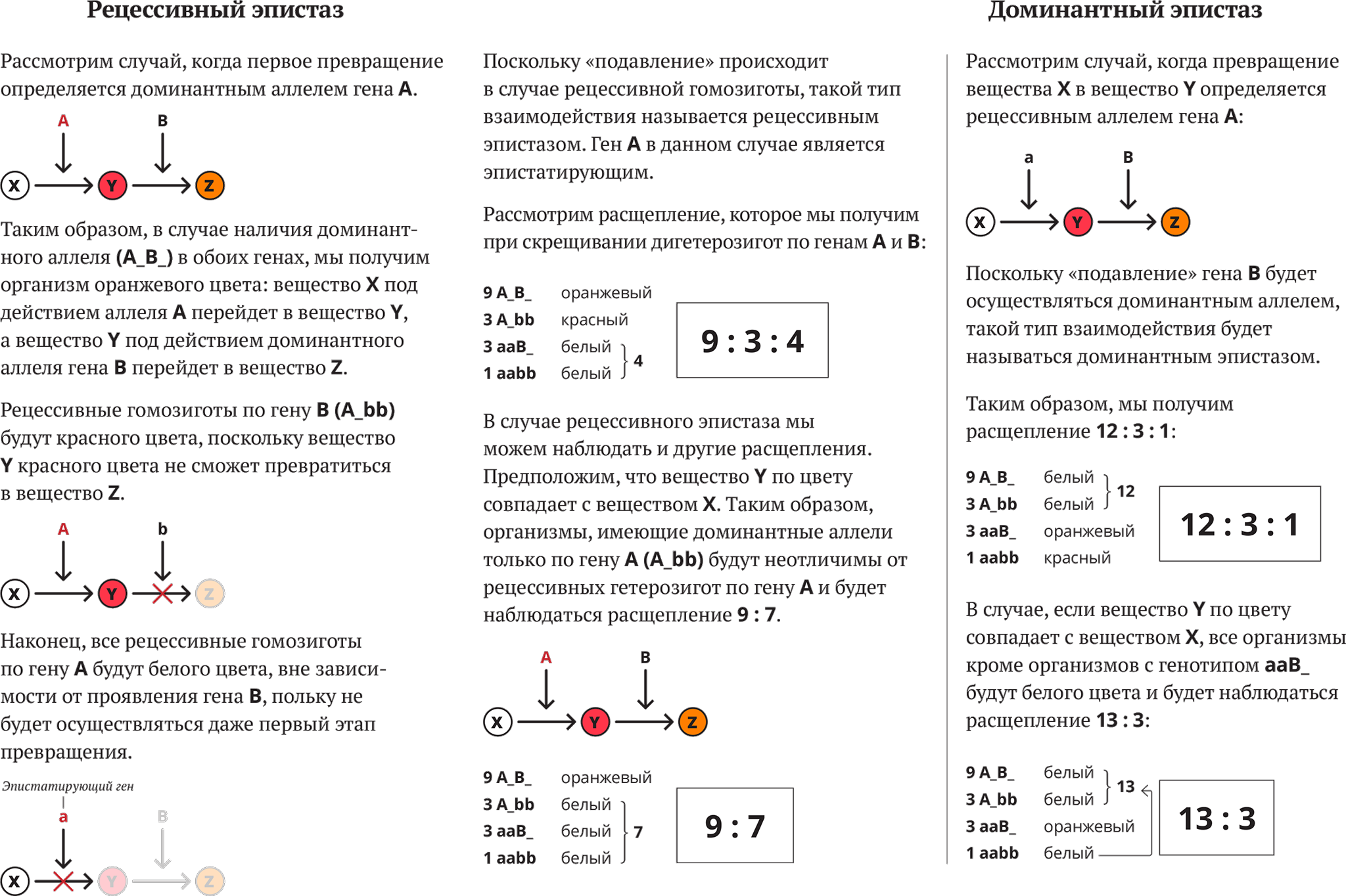
Эпистаз принято делить на два типа: доминантный и рецессивный. Под доминантным эпистазом понимают подавление одним доминантным геном действия другого гена.
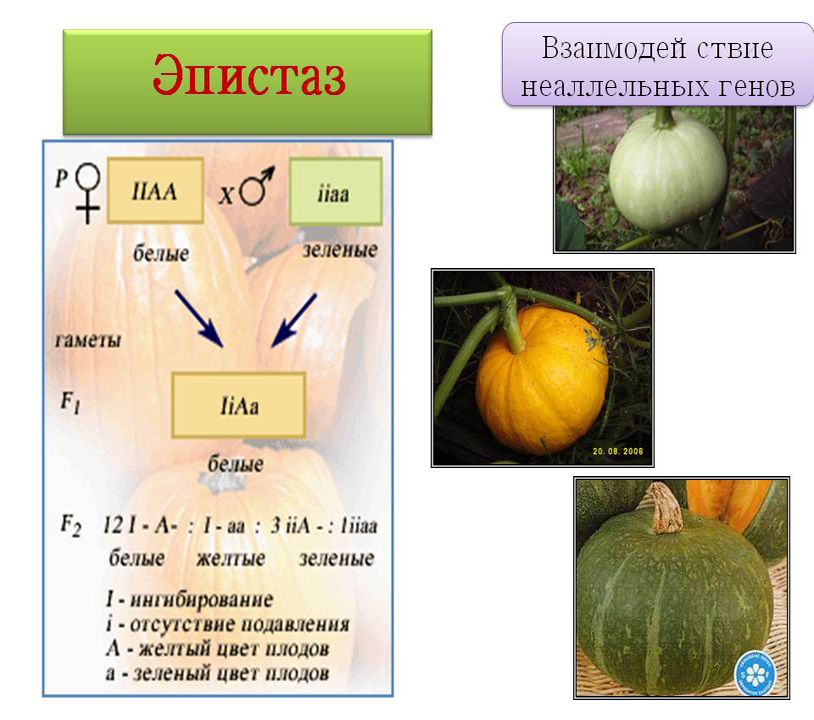
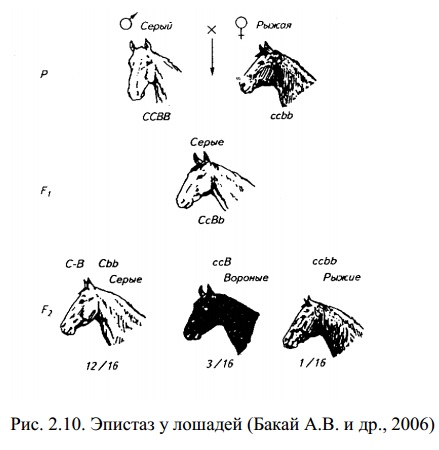
Окраска шерсти у мышей контролируется парой генов, находящихся в разных локусах.
Эпистатический ген определяет наличие окраски и имеет два аллеля: доминантный, определяющий окрашенную шерсть, и рецессивный, обусловливающий альбинизм (белая окраска). Гипостатический ген определяет характер окраски и имеет два аллеля: агути (доминантный, определяющий серую окраску) и черный (рецессивный). Мыши могут иметь серую или черную окраску в зависимости от своих генотипов, но наличие окраски возможно только в том случае, если у них одновременно имеется аллель окрашенной шерсти.
Мыши, гомозиготные по рецессивному аллелю альбинизма, будут альбиносами даже при наличии у них аллелей агути и черной шерсти. Возможны три разных фенотипа: агути, черная шерсть и альбинизм. При скрещивании можно получить эти фенотипы в различных соотношениях в зависимости от генотипов скрещиваемых особей.
Примеры эпистаза
Так, рецессивные гены окраски собак из локуса С не позволяют синтезироваться пигментам, определяющим окраску шерсти.
Собака гомозиготная по ним — белая.
Такой тип эпистаза называется рецессивным. Скрещивание белой черноносой собаки с коричневой приведет к рождению в первом поколении гетерозиготных черных собак, а скрещивание гибридов F, между собой — к рождению 9 черных, 3 коричневых и 4 белых, т. е. будет наблюдаться отклонение от нормального расщепления 9:3:3:1.
Скрещивание белой кошки, имеющей доминантный белый окрас, обусловленный доминантным аллелем W, с черной, напротив, приведет к рождению в первом поколении белых гетерозиготных котят.
Расщепление в F2 даст вместо ожидаемых 9:3:3:1 соотношение 12:3:1, где 12 — белых, 3 — полосатых и 1 черный котенок. Такой тип эпистаза называется доминантным. Отсутствие необходимых данных о роли первичных продуктов многих генов (ферментов, пептидов) в формировании сложных признаков, часто не позволяет точно установить характер взаимодействия неаллельных локусов, участвующих в биохимических процессах и составляющих основу образования этих признаков.
В одних случаях развитие признака при наличии двух неаллельных генов в доминантном состоянии рассматривают как комплементарное воздействие, в других — неразвитие признака, определяемого одним из генов при отсутствии другого гена в доминантом состоянии, расценивают как рецессивный эпистаз; если же признак развивается при отсутствии доминантного аллеля неаллельного гена, а в его присутствии не развивается, говорят о доминантом эпистазе.
Что такое эпистаз и криптомерия?
При неаллельном взаимодействии одна пара аллелей может подавлять проявление другой. Такой тип взаимодействия называется эпистазом, а ген, оказывающий ингибирующее действие, — ингибитором, супрессором или подавителем.
Так, скрещивание лука с красными и белыми луковицами дает в F1 растения только с белыми луковицами, а в F2 — расщепление: 12/16 — с белыми, 3/16 — с красными и 1/16 — с желтыми луковицами.
Обозначив ген, определяющий красный цвет луковиц, через Y, ген, определяющий желтый цвет, — через y, ген-ингибитор — через I, его рецессивный аллеломорф, не оказывающий подавляющего действия, — через i, проведем скрещивание:

Расщепление по фенотипу 12:3:1.
Таким образом, формы, в генотипе которых присутствовал ген-ингибитор, оказывались с белыми луковицами (доминантный эпистаз I подавляет как Y, так и y). Нужно отметить, что гетерозиготы по I-i паре имеют слабую окраску вокруг шейки луковицы, в то время как формы II совершенно белые.
Несколько иной характер наследований наблюдается при скрещивании растений лука, если рецессивный ген с (colorless — бесцветный) в гомозиготном состоянии определяет образование белых луковиц, а доминантный С — окрашенных. Рецессивный эпистаз ее подавляет как Y, так и y:

Расщепление по фенотипу 9:3:4.
Этот случай интересен в практическом отношении. Дело в том, что окрашенные луковицы устойчивы к пятнистости (болезнь лука), которая является результатом присутствия определенных веществ, связанных с образованием пигмента. Рецессивные белые формы ссyy, лишенные пигмента, подвержены болезни, а белые формы с генотипом ccY- устойчивы к ней.






Скрещивание некоторых белых пород кур давало в F1 только белых птиц, а в F1 наряду с белыми появлялись и окрашенные (в отношении 13:3). Очевидно, одна из скрещиваемых форм потенциально способна синтезировать пигмент, но его синтез блокируется наличием в генотипе гена-ингибитора. Генотип такой формы можно представить как I-C- (С- определяет наличие предшественника пигмента; I — ингибитор):

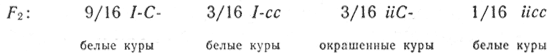
Расщепление по фенотипу 13:3.
В ряде случаев отдельные гены не проявляют своего действия фенотипически, если не взаимодействуют с другими неаллельными им генами. Такой тип взаимодействия называется криптомерией, а ген, присутствие которого в генотипе необходимо для фенотипического проявления признака, можно назвать геном-проявителем. Так, у льна В — ген, определяющий голубую окраску цветков, b — розовую. Однако для проявления окраски необходимо наличие в генотипе доминантного гена (проявителя) А. Его отсутствие в генотипе определяет белую окраску цветков:

Расщепление по фенотипу 9:3:4.
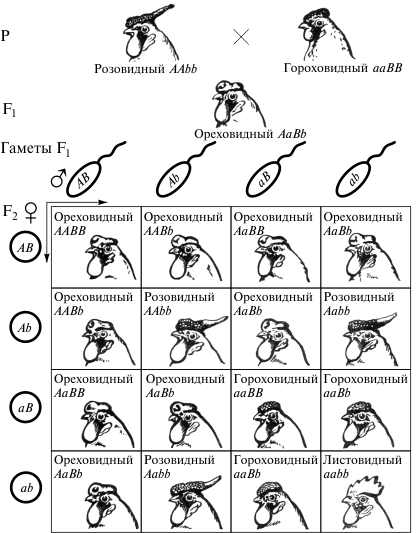
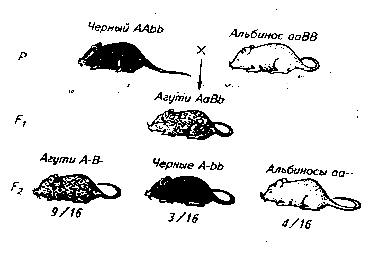
У грызунов (кроликов, белок, морских свинок, мышей и др.) окраска дикого типа — агути зависит от действия двух генов: гена, определяющего ее развитие (С), и гена, отвечающего за распределение пигмента по длине волоса (А — зональное распределение пигмента; а — равномерное его распределение).
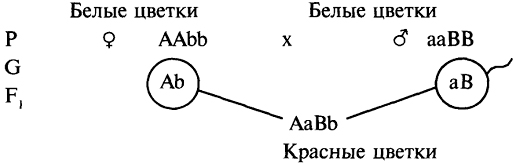
При скрещивании черных мышей с белыми (альбиносами) все гибриды в первом поколении оказываются агути, а в F2 наблюдается расщепление (9/16 составляют агути, 3/16 — черные и 4/16 — белые мыши):

Расщепления в примерах на первый взгляд кажутся несовпадающими с менделеевским расщеплением, но при более глубоком анализе выявляется их общая основа. Для знания того, к какому типу наследования принадлежит каждый конкретный случай, необходимо выяснить биохимическую основу данного признака. Незнание этой биохимической основы приводит к положению, при котором один и тот же случай может быть отнесен к различным типам взаимодействия генов (в частности, пример с наследованием окраски у мышей может рассматриваться как обычное комплементарное наследование).
Эпистатическое действие генов
Окраска агути (рыжевато-серая) определяется доминантным геном А, его рецессивный аллель а в гомозиготном состоянии определяет черную окраску. Доминантный ген другой нары С определяет развитие пигмента, а гомозиготы по его рецессивному аллелю с являются альбиносами (отсутствие пигмента в шерсти и радужной оболочке глаз).
Особи, являющиеся доминантными гомозиготами по гену С или гетерозиготные по этому гену, будут иметь окраску шерсти в соответствии со своим генотипом по гену А (например, ААСС и ААСс будут рыжевато-серые агути, ааСС и ааСс — черные). Но все особи, гомозиготные по рецессивному гену-супреесору (ААсс, Аасс и аасс), будут альбиносами.
Т.Л. Богданова «Пособие для поступающих в вузы»
Как известно, доминирование есть подавление действия одной аллели другой аллелью, представляющих собой один ген: А > а, В > b, С > с и т. д.
Но существует взаимодействие, при котором аллель одного из генов подавляет действие аллелей других генов, например А > В или В А, а > В или b > А и т. д. Такое явление «доминирования» между генами называется эпистазом.
Эпистатическое взаимодействие генов по своему характеру противоположно комплементарному взаимодействию.
Гены, подавляющие действие других генов, называются супрессорами, или ингибиторами.
Они могут быть как доминантными, так и рецессивными. Гены-супрессоры известны у животных (млекопитающие, птицы, насекомые) и у растений. Обычно они обозначаются I или Su в случае доминантного состояния генов и i или su для их рецессивных аллелей (от английских слов inhibitor или supressor).
В настоящее время эпистаз делят на два типа: доминантный и рецессивный.
Под доминантным эпистазом понимают подавление доминантной аллелью одного гена действия аллельной пары другого гена.
Из многих примеров доминантного эпистаза, установленных как на животных, так и на растениях, приведем лишь отдельные.
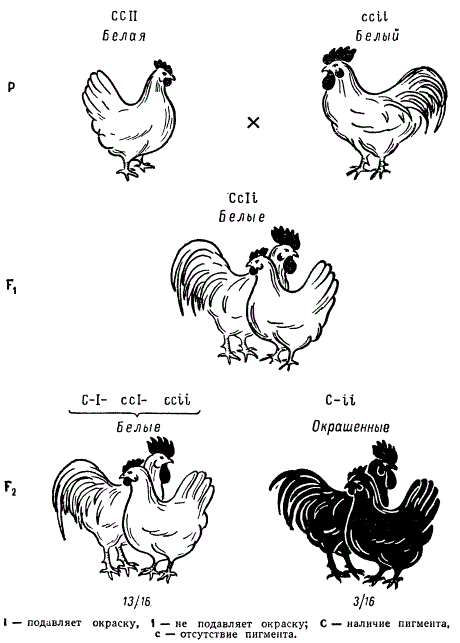
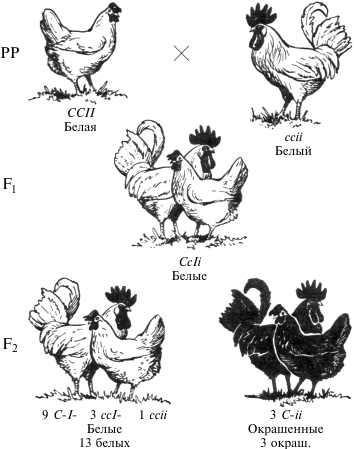
Некоторые породы кур имеют белое оперение (белый леггорн, белый примутрок и др.), другие же породы имеют окрашенное оперение (австралорп, ньюгемпшир, полосатый плимутрок и др.). Белое оперение разных пород кур определяется несколькими различными генами.
![Что такое эпистаз и криптомерия? [1989 морозов е.и., тарасевич е.и., анохина в.с. - генетика в вопросах и ответах]](https://rusinfo.info/wp-content/uploads/8/1/5/815ec032fa70e7ea0376ff23b1b40339.jpg)




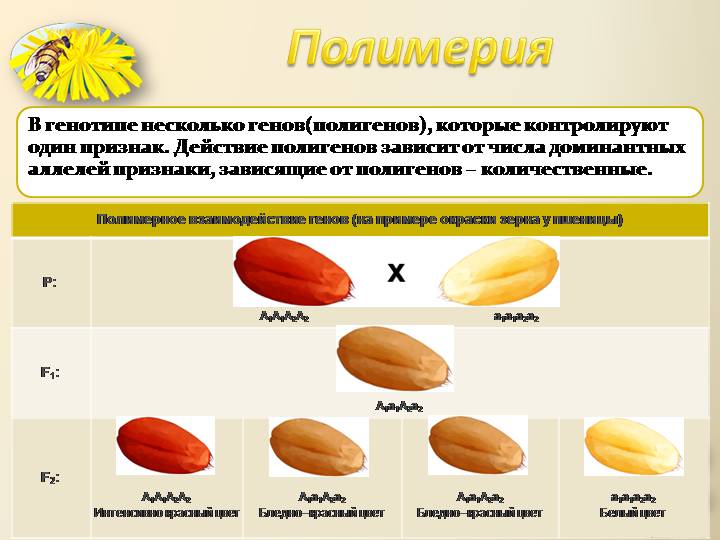
Так, например, доминантная белая окраска определяется генами CCII (белые леггорны), а рецессивная белая — ccii (белые суссексы, белые минорки, белые плимутроки). Ген С определяет наличие предшественник пигменту (хромогена), т. е. окрашенность пера, его аллель с — отсутствие хромогена и, следовательно, неокрашенность пера птицы. Ген I является подавителем действия гена С, аллель i не подавляет его действия. В присутствии даже одной дозы гена I в генотипе птицы действие генов окраски не проявится.
Поэтому при скрещивании белых леггорнов CCII с цветными породами CCii, как правило, доминирует белая окраска CCIi. При скрещивании белых плимутроков ccii с окрашенными породами CCii гибриды оказываются окрашенными Ccii. Следовательно, у леггорнов белая окраска является доминантной, а у плимутроков — рецессивной.
Если же производится скрещивание белых леггорнов CCII с рецессивными белыми плимутроками ccii, то в первом поколении цыплята оказываются тоже белыми CcIi.
При скрещивании между Собой гибридов F1 во втором поколении имеет место расщепление по окраске в отношении 13/16 белых : 3/16 окрашенных.
Наследование окраски у кур при взаимодействии двух пар генов (эпистаз)
Как можно объяснить полученное отношение?
Прежде всего, данное отношение говорит о расщеплении по двум генам, это расщепление можно представить как 9(С—I—) + 3(ccI—) + 1(ccii) = 13 и 3(С—ii), что соответствует формуле 9:3:3:1.
Очевидно, в этом случае окраска леггорнов обязана не присутствию особых генов белой окраски, а действию гена — подавителя окраски (I—). Тогда генотип гомозиготных белых леггорнов должен быть ССII, где I является геном — подавителем окраски, а С — геном окраски.
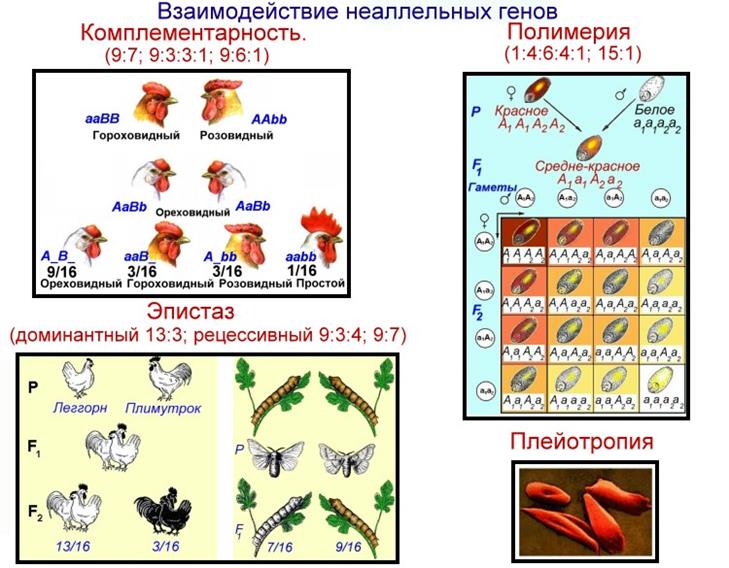
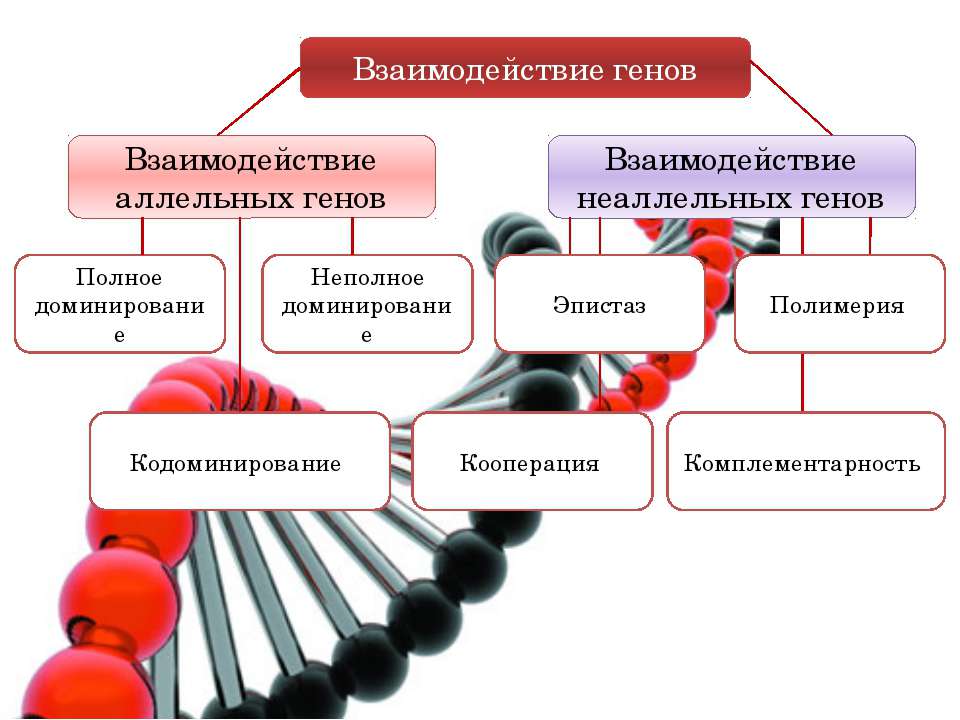
Полимерия
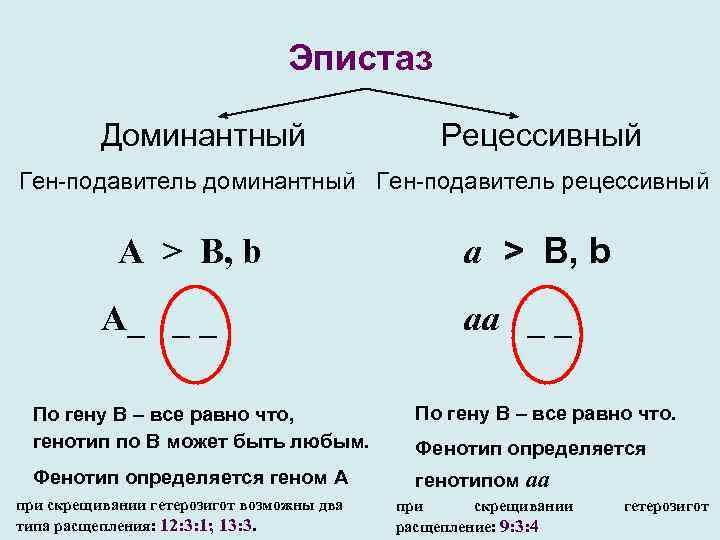
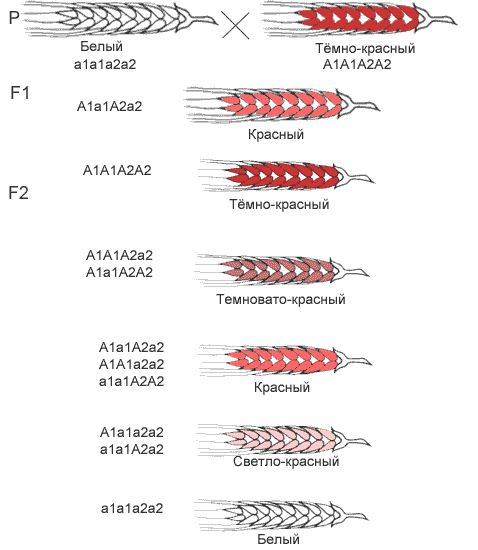
Это вид взаимодействия двух и более пар неаллельных генов, доминантные аллели которых однозначно влияют на развитие одного и того же признака. Полимерное действие генов может быть кумулятивным и некумулятивным. При кумулятивной полимерии интенсивность значения признака зависит от суммирующего действия генов: чем больше доминантных аллелей, тем больше степень выраженности признака. При некумулятивной полимерии количество доминантных аллелей на степень выраженности признака не влияет, и признак проявляется при наличии хотя бы одного из доминантных аллелей. Полимерные гены обозначаются одной буквой, аллели одного локуса имеют одинаковый цифровой индекс, например А1а1А2а2А3а3.
Кумулятивная полимерия имеет место при наследовании окраски зерновок пшеницы, чешуек семян овса, роста и цвета кожи человека и т.д.
| P | A1A1A2A2черные чешуйки | × | a1a1a2a2белые чешуйки |
| Типы гамет | A1A2 | a1a2 | |
| F1 | A1a1A2a2серые чешуйки, 100% | ||
| P | A1a1A2a2серые чешуйки | × | A1a1A2a2серые чешуйки |
| Типы гамет | A1A2 A1a2 a1A2 a1a2 | A1A2 A1a2 a1A2 a1a2 |
| A1A2 | A1a2 | a1A2 | a1a2 | ||
| A1A2 | A1A1A2A2черные | A1A1A1a2темно-серые | A1a1A2A2темно-серые | A1a1A2a2серые | |
| A1a2 | A1A1A2a2темно-серые | A1A1a2a2серые | A1a1A2a2серые | A1a1а2a2светло-серые | |
| a1A2 | A1a1A2A2темно-серые | A1a1A2a2серые | a1a1A2A2серые | a1a1A2a2светло-серые | |
| a1a2 | A1a1A2a2серые | A1a1a2a2светло-серые | a1a1A2a2светло-серые | a1a1a2a2желтые |
Черные чешуйки семян у овса — 1/16, темно-серые чешуйки семян у овса — 4/16, серые чешуйки семян у овса — 6/16, светло-серые чешуйки семян у овса — 4/16, желтые чешуйки семян у овса — 1/16. Расщепление по фенотипу 1:4:6:4:1.
Некумулятивная полимерия имеет место при наследовании формы плодов пастушьей сумки.
| P | A1A1A2A2треугольные | × | a1a1a2a2овальные |
| Типы гамет | A1A2 | a1a2 | |
| F1 | A1a1A2a2треугольные, 100% | ||
| P | A1a1A2a2треугольные | × | A1a1A2a2треугольные |
| Типы гамет | A1A2 A1a2 a1A2 a1a2 | A1A2 A1a2 a1A2 a1a2 |
| A1A2 | A1a2 | a1A2 | a1a2 | ||
| A1A2 | A1A1A2A2треугольные | A1A1A1a2треугольные | A1a1A2A2треугольные | A1a1A2a2треугольные | |
| A1a2 | A1A1A2a2треугольные | A1A1a2a2треугольные | A1a1A2a2треугольные | A1a1а2a2треугольные | |
| a1A2 | A1a1A2A2треугольные | A1a1A2a2треугольные | a1a1A2A2треугольные | a1a1A2a2треугольные | |
| a1a2 | A1a1A2a2треугольные | A1a1a2a2треугольные | a1a1A2a2треугольные | a1a1a2a2овальные |
Треугольная форма плодов у пастушьей сумки — 15/16, овальная форма плодов у пастушьей сумки — 1/16.
Плейотропия — множественное действие генов. Плейотропное действие генов имеет биохимическую природу: один белок-фермент, образующийся под контролем одного гена, определяет не только развитие данного признака, но и воздействует на вторичные реакции биосинтеза других признаков и свойств, вызывая их изменение.
Плейотропное действие генов впервые было обнаружено Г. Менделем, который установил, что у растений с пурпурными цветками всегда имелись красные пятна в пазухах листьев, а семенная кожура была серого или бурого цвета. То есть развитие этих признаков определяется действием одного наследственного фактора (гена).
У человека встречается рецессивная наследственная болезнь — серповидно-клеточная анемия. Первичным дефектом этой болезни является замена одной из аминокислот в молекуле гемоглобина, что приводит к изменению формы эритроцитов. Одновременно с этим возникают нарушения в сердечно-сосудистой, нервной, пищеварительной, выделительной системах. Это приводит к тому, что гомозиготный по этому заболеванию ребенок погибает в детстве. Причиной синдрома Марфана является доминантная мутация гена, контролирующего одновременно рост, длину пальцев, формирование интеллекта и форму хрусталика. Для человека с этим синдромом характерен комплекс следующих признаков — высокий рост, очень длинные гибкие («паучьи») пальцы, повышенный интеллект, близорукость.
Плейотропия широко распространена. Изучение действия генов показало, что плейотропным эффектом, очевидно, обладают многие, если не все, гены.
Таким образом, выражение «ген определяет развитие признака» в значительной степени условно, так как действие гена зависит от других генов — от генотипической среды. На проявление действия генов влияют и условия окружающей внешней среды. Следовательно, генотип является системой взаимодействующих генов.
-
Перейти к лекции №19 «Генетика пола»
-
Перейти к лекции №21 «Изменчивость»
-
Смотреть оглавление (лекции №1-25)
Классическое фенотипическое соотношение 9: 3: 3: 1
Эта пропорция возникает из-за комбинации анализа наследования двух разных символов. То есть это продукт сочетания двух независимых фенотипических сегрегаций (3: 1) X (3: 1).








Когда Мендель, например, анализировал высоту растения или цвет семени, каждый символ был разделен от 3 до 1. При анализе в целом, даже если они были двумя разными символами, каждый из них был отделен от 3 до 1. То есть они были распределены независимо.
Однако, когда Мендель проанализировал символы в парах, они привели к известным фенотипическим классам 9, 3, 3 и 1. Но эти классы были суммами из двух символов другой. И никогда, ни один персонаж не влиял на то, как проявился другой.

Механизмы эпистаза генов
Гены – супрессоры обладают способностью подавлять действие других генов, гены ингибиторы оказывают аналогичное действие. Реакция, возникающая от такого взаимодействия, может проявиться в рецессивном и доминантном вариантах.
Доминантный эпистаз наблюдается в том случае, если доминантная аллель гена – супрессора подавляет действие другого гена. Такая аллель, как правило, обозначается буквой I.
В свою очередь рецессивный эпистаз наблюдается в диаметрально противоположном случае при угнетении одного гена другим.
Фенотипическое расщепление при каждом виде эпистаза существенно различается. При доминантном эпистазе в случае скрещивания дигетерозигот расщепление может быть в следующих соотношениях:
- тринадцать к трем и семь к шести к трем;
- двенадцать к трем и к одному.
При рецессивном эпистазе расщепление может быть в следующих соотношениях:
- девять к трем и к четырём;
- тринадцать к трем.
Взаимодействие по типу эпистаза противоположно комплементарному. При наличии комплиментарного взаимодействия наблюдается дополнение одного гена другим. Эпистаз очень схож с доминированием, но разница проявляется в участие аллеля, принадлежащего к аллеморфной паре.
При эпистазе аллель одного гена подавляет проявление аллеля из другой аллеломорфной пары, т. е. неаллельного гена. Фенотипически эпистаз выражается в отклонении от расщепления, которое ожидается от стандартного наследования. Следует отметить тот факт, что нарушения законов Менделя в данном случае не происходит, поскольку распределения аллелей всех взаимодействующих генов соответствует закону независимого комбинирования тех или иных признаков.
Эпистатическая система наследования обнаружена у кур. Некоторые породы кур имеют белое оперение (плимутрок, леггорн), другие породы имеют окрашенное оперение (полосатый плимутрок). При этом белое оперение различных пород кур определяется несколькими различными генами. Например, белая доминантная окраска определяется генами CCII, а рецессивная белая – ccii. Возможно и другое условное обозначение.
При этом ген С позволяет определить наличие предшественника пигмента или исследовать окрашенность пера и гены, отвечающие за не окрашенность пера. При скрещивании между собой гибридов F1 во втором поколении имеет место расщепление по окраске в отношении 13 белых: 3 окрашенных.
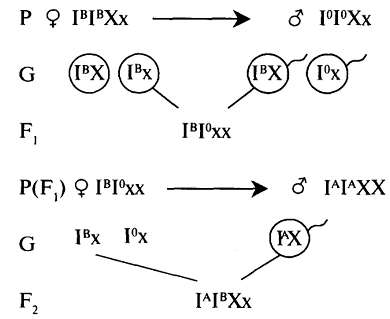
Также можно привести и другие примеры эпистатического взаимодействия генов. У человека существует так называемый бомбейский феномен. Люди, которые гетерозиготные по гену FUTI обладают нарушенной экспрессией генов, которые кодируют ферменты, отвечающие за образование эритроцитов A и B. На эритроцитах вместо нормальных генов образуются дефектные гены. Эти люди имеют группу крови O независимо от аллелей генов, которые определяют тип группы крови по системе ABO, не имеют антиегна A, антигена B.
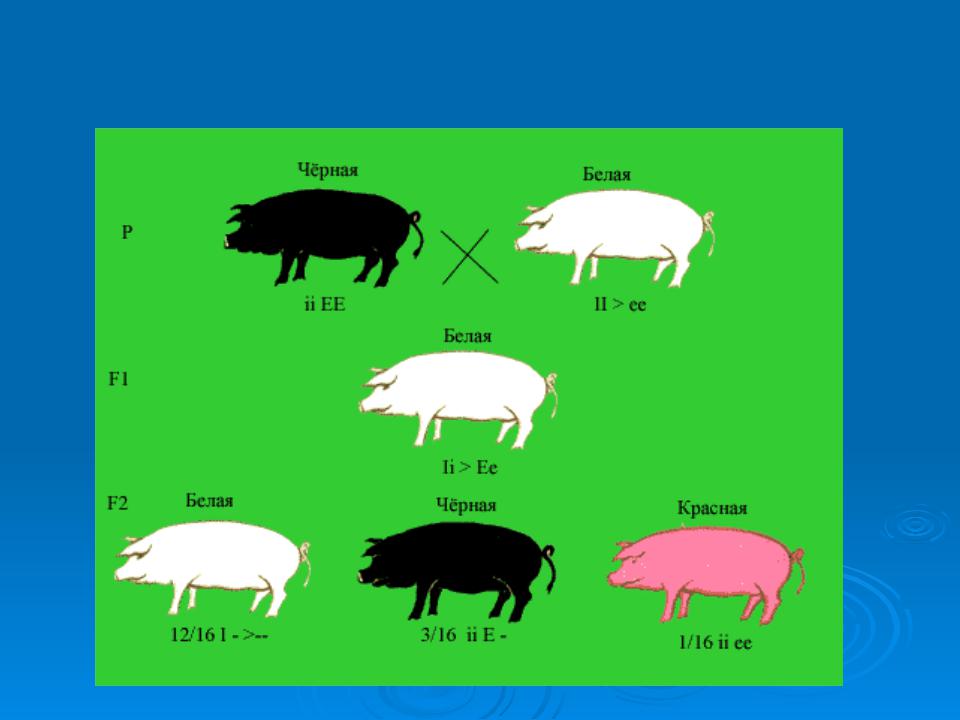
Гены, которые определяют окраску шерсти у мышей также обладают эпистатическим взаимодействием. Окраска агаути у мышей обусловлена чередованием кольцевых пигментных полос по длине каждой части волосяного покрова. Она доминирует над чёрной окраской и определяется рецессивным мутантным аллелем. Если рецессивная мутация имеется в другом локусе, то пигментация полностью исчезает, и эта ситуация совершенно не зависит от того, какими аллелями представлен ген A.
Доминантный эпистаз у тыквы проявляется в окраске плодов. Доминантный аллель тыквы обуславливает ее белую окраску, вне зависимости от того, какими аллелями представлен ген B. Если локус A представлен рецессивной аллелью, то растения с генотипами BB и Bb имеют жёлтые плоды, а bb — зелёные.
Встречаются ситуации, когда наблюдается двойной эпистаз. Он встречается в тех случаях, когда встречаются гены, кодирующие ферменты одного метаболического пути. Если внутри клетки не будет создаваться нормальная форма одного из ферментов, то также не будет образовываться промежуточный продукт, который является фундаментом для всех последующих стадий. В результате нормативного продукта не будет образовываться при любой комбинации аллелей.
Также в качестве примера можно привести гены мухи – дрозофилы, которые кодируют ферменты – катализаторы, участвующие в превращении триптофана, который приобретает бурую окраску во время химических превращений. Если в процессе будет участвовать только рецессивный ген, то фенотип мух по окраске будет существенно отличаться от дикого типа.
Таким образом, эпистатическое взаимодействие неаллельных генов является одним из примеров подавления одного гена другим. Он позволяет отследить закономерности наследования генов и отразить специфику их взаимодействия внутри генотипа.
Полное доминирование
Полное доминирование — это вид взаимодействия аллельных генов, при котором фенотип гетерозигот не отличается от фенотипа гомозигот по доминанте, то есть в фенотипе гетерозигот присутствует продукт доминантного гена. Полное доминирование широко распространено в природе, имеет место при наследовании, например, окраски и формы семян гороха, цвета глаз и цвета волос у человека, резус-антигена и мн. др.
Наличие резус-антигена (резус-фактора) эритроцитов обусловливается доминантным геном Rh. То есть генотип резус-положительного человека может быть двух видов: или RhRh, или Rhrh; генотип резус-отрицательного человека — rhrh. Если, например, мать — резус-отрицательная, а отец резус-положительный и гетерозиготен по этому признаку, то при данном типе брака с одинаковой вероятностью может родиться как резус-положительный, так и резус-отрицательный ребенок.
| Р | rhrhрезус-отрицательная | × | Rhrhрезус-положительный |
| Типы гамет | rh | Rh rh | |
| F | Rhrhрезус-положительный50% | rhrhрезус-отрицательный50% |
Между резус-положительным плодом и резус-отрицательной матерью может возникнуть резус-конфликт.
ссылки
- Брукер Р.Дж. (2017). Генетика: анализ и принципы. McGraw-Hill Higher Education, Нью-Йорк, Нью-Йорк, США.
- Goodenough, U. W. (1984) Genetics. W. B. Saunders Co. Ltd, Пкиладелия, Пенсильвания, США.
- Griffiths A.J.F., Wessler R., Carroll S.B., Doebley J. (2015). Введение в генетический анализ (11-е изд.). Нью-Йорк: У. Х. Фриман, Нью-Йорк, Нью-Йорк, США.
- Miko, I. (2008) Epistasis: взаимодействие генов и фенотипические эффекты. Природоведческое образование 1: 197. nature.com
- Уайт Д., Рабаго-Смит М. (2011). Генотип-фенотипические ассоциации и цвет глаз человека. Журнал генетики человека, 56: 5-7.
- .
Явление эпистаза генов
Необходимо ответить тот факт, что очень редко развитие того или иного признака определяется одним геном. В большинстве случаев какой – либо ген в единственном экземпляре становится фундаментом для развития различных признаков. Такое явление называется плейотропным взаимодействием генов. Также весьма распространено такое явление, когда один признак контролируется несколькими генами.
![Что такое эпистаз и криптомерия? [1989 морозов е.и., тарасевич е.и., анохина в.с. - генетика в вопросах и ответах]](https://rusinfo.info/wp-content/uploads/9/e/c/9ec50decfa513195f25204364995841b.png)






Определение 2
Неаллельные гены – это гены, которые расположены в различных местах (участках хромосом) и кодируют разные белки.
Неаллельные гены могут взаимодействовать между собой, когда один признак проявляется под действием нескольких генов.
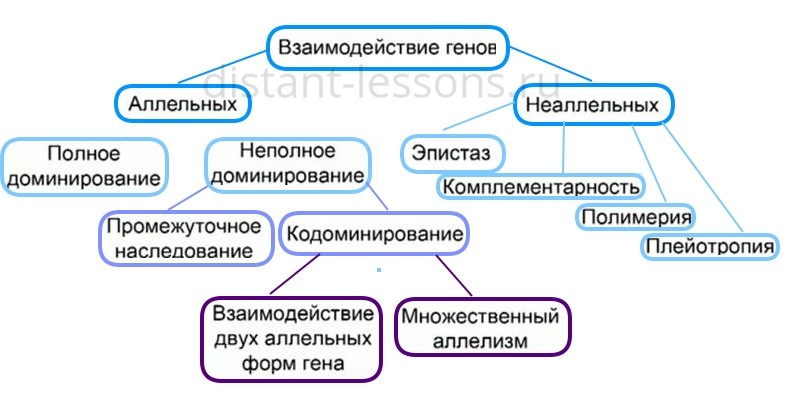
В науке выделяется три формы взаимодействия генов между собой:
- комплементарность (или такое взаимодействие генов, которые дополняют друг друга и обуславливают развитие нового признака);
- полимерия или процесс генетического взаимодействия, при котором неаллельные гены влияют в одном напралвении на развитие одинакового признака;
- эпистаз (подавление гена иным геном).
Все указанные типы неаллельных взаимодействий встречаются с равной степенью вероятности и должны оцениваться с позиций гибридологического подхода.
Другие эпистатические фенотипические пропорции
Из доли, предложенной во втором законе Менделя, у нас есть другие дополнительные случаи, которые заслуживают упоминания.
При измененном соотношении 9: 4: 3 мы называем это рецессивным эпистазом по уважительной причине. Когда ген является гомозиготным по рецессивному гену, он избегает экспрессии другого гена, даже если он является доминантным.
Возьмем в качестве примера рецессивный эпистаз генотипа аа о гене В. Класс 9 — это уже признанный 9 A_B_. Для класса 4 к классу 1 aabb они должны быть добавлены с тем же фенотипом, что и для класса 3 aaB_. Класс 3 будет классом 3 A_bb.
При эпистатическом взаимодействии дуплицированных генов наблюдается фенотипическое соотношение 9: 6: 1. Все особи класса 9 A_B_ имеют по крайней мере один аллель каждого гена или В. Все имеют одинаковый фенотип.
Напротив, в классах 3 A_bb и 3 aaBb присутствуют только доминантные аллели либо A, либо B. В этом случае также существует один и тот же фенотип, но отличающийся от других. Наконец, в 1-м классе aabb доминантный аллель ни одного из генов отсутствует и представляет другой фенотип.
Возможно, класс, который больше всего смущает, — это доминирующий эпистаз, который показывает фенотипическую пропорцию 12: 3: 1. Здесь доминирование A (эпистатическое) над B (гипостатическое) заставляет класс 9 A-B_ присоединиться к классу 3 A_bb.
Фенотип B проявится только тогда, когда A не присутствует в классе 3 aaB_. Двойной рецессивный класс 1 aabb не будет проявлять фенотип, связанный с геном A / A ни с геном Б / б.
Другими эпистатическими фенотипическими пропорциями, которые не имеют конкретного названия, являются 7: 6: 3, 3: 6: 3: 4 и 11: 5.
Методы и модельные системы
Регрессионный анализ
Количественная генетика фокусируется на генетической изменчивости из-за генетических взаимодействий. Любые два локусных взаимодействия с определенной частотой гена можно разложить на восемь независимых генетических эффектов с помощью взвешенной регрессии . В этой регрессии наблюдаемые генетические эффекты двух локусов рассматриваются как зависимые переменные, а «чистые» генетические эффекты используются как независимые переменные. Поскольку регрессия является взвешенной, распределение между компонентами дисперсии будет изменяться в зависимости от частоты генов. По аналогии можно расширить эту систему до трех или более локусов или до цитоядерных взаимодействий.
Двойные мутантные циклы
При анализе эпистаза в гене можно использовать сайт-направленный мутагенез для генерации различных генов, а их белковые продукты могут быть проанализированы (например, на стабильность или каталитическую активность). Иногда это называют двойным мутантным циклом, и он включает производство и анализ белка дикого типа, двух одиночных мутантов и двойного мутанта. Эпистаз измеряется как разница между совокупными эффектами мутаций и суммой их индивидуальных эффектов. Это можно выразить как свободную энергию взаимодействия. Ту же методологию можно использовать для исследования взаимодействий между более крупными наборами мутаций, но все комбинации должны быть получены и проанализированы. Например, существует 120 различных комбинаций из 5 мутаций, некоторые или все из которых могут проявлять эпистаз …
Вычислительный прогноз
Были разработаны многочисленные вычислительные методы для обнаружения и характеристики эпистаза. Многие из них полагаются на машинное обучение для обнаружения неаддитивных эффектов, которые могут быть упущены статистическими подходами, такими как линейная регрессия. Например, многофакторное снижение размерности (MDR) было разработано специально для непараметрического и безмодельного обнаружения комбинаций генетических вариантов, которые предсказывают фенотип, такой как статус заболевания, в популяциях людей . . Некоторые из этих подходов широко рассмотрены в литературе. Еще совсем недавно было показано , что методы, использующие идеи теоретической информатики ( преобразование Адамара ) или вывод максимального правдоподобия, позволяют отличать эпистатические эффекты от общей нелинейности в структуре карты генотип-фенотип, в то время как другие использовали анализ выживаемости пациентов для выявления нестандартных факторов. линейность.
ЭПИСТАЗ. ТИПЫ ЭПИСТАЗА. ХАРАКТЕР РАСЩЕПЛЕНИЯ ПРИЗНАКОВ. ПРИМЕРЫ.
Белые плимутроки по генотипу должны быть гомозиготными по двум рецессивным факторам ccii, где с — отсутствие окраски и i — отсутствие подавления окраски. В силу эпистатирования I > С гибридные куры первого поколения CcIi должны быть белыми. В F2 все куры с генотипами 9/16 С—I—, 3/16 ccI— и 1/16 ccii также должны быть белыми, и лишь куры одного фенотипического класса 3/16 (С—ii) оказываются окрашенными, поскольку здесь содержится ген окраски и нет его подавителя.
Таким образом, подавление действия доминантной аллели гена, Определяющего развитие окраски, доминантной аллелью другого гена (подавителем) обусловливает в F2 расщепление по фенотипу в отношении 13 : 3.
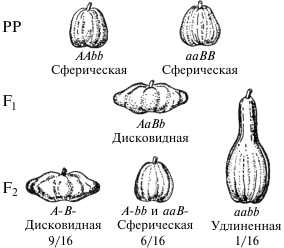
Доминантный эпистаз может давать и другое отношение при расщеплении в F2 по фенотипу, а именно 12 : 3 : 1.
В этом случае форма гомозиготная по обоим рецессивным факторам aabb, будет фенотипически отличима от форм с доминантными аллелями двух генов А—В— и форм с одной из них: ааВ— и А—bb.
Такое расцепление установлено для наследования окраски плодов у тыквы, кожуры у лука и других признаков. В этом случае в расщеплении также принимает участие доминантный ингибитор.




![Что такое эпистаз и криптомерия? [1989 морозов е.и., тарасевич е.и., анохина в.с. - генетика в вопросах и ответах]](https://rusinfo.info/wp-content/uploads/3/a/1/3a1efa98e2cbd1506bac4adbd4a5df74.jpg)

Мы разобрали взаимодействие только двух генов. В действительности по типу эпистаза взаимодействуют многие гены. Гены-подавители обычно не определяют сами какой-либо качественной реакции или синтетического процесса, а лишь подавляют действие других генов.
Однако когда мы говорим, что ген-подавитель не имеет своего качественного влияния на признак, то это относится только к данному признаку. На самом же деле ингибитор, подавляя, например, пигментообразование, может оказывать плейотропное действие на другие свойства и признаки.
Под рецессивным эпистазом понимают такой тип взаимодействия, когда рецессивная аллель одного гена, будучи в гомозиготном состоянии, не дает возможности проявиться доминантной или рецессивной аллелям других генов: аа > В или aa > bb.
Мы уже имели случай познакомиться с расщеплением 9:3:4 как результатом комплементарного взаимодействия генов.
Но эти же случаи можно рассматривать и как пример рецессивного эпистаза.
При скрещивании черных мышей (ААbb) с белыми (ааВВ) все особи F1 (АаВb) имеют окраску типа агути, а в F29/16 всех особей сказываются агути (А—В—), 3/16 черные (A—bb) и 4/16 белые (ааВ— и aabb).
Эти результаты можно объяснить, предположив, что имеет место рецессивный эпистаз типа ааВ—. При этом мыши генотипа ааВ— оказываются белыми потому, что ген а в гомозиготном состоянии, обусловливая отсутствие пигмента, препятствует тем самым проявлению гена-распределителя пигмента В.
Кроме описанных случаев одинарного рецессивного эпистаза, действуют и такие, когда рецессивная аллель каждого из генов в гомозиготном состоянии одновременно реципрокно подавляет действием доминантных аллелей каждого из генов, т.
е. аа эпистатирует над В—, a bb над А—. Такое взаимодействие двух подавляющих рецессивных генов называют двойным рецессивным эпистазом. При этом в дигибридном скрещивании расщепление по фенотипу будет соответствовать 9 : 7, как и в случае комплементарного взаимодействия генов.
Следовательно, одно и то же отношение можно трактовать и как комплементарное взаимодействие, и как эпистатирование.
Сам по себе генетический анализ наследования при взаимодействии генов без учета биохимии и физиологии развития признака в онтогенезе не может раскрыть природы этого взаимодействия. Но без генетического анализа нельзя понять наследственной детерминации развития этих признаков.