Результаты гаструляции.
Конечный результат гаструляции заключается в образовании двуслойного зародыша. Наружный слой зародыша (эктодерма) образован мелкими, часто – пигментированными клетками, не содержащими желтка; из эктодермы в дальнейшем развиваются такие ткани, как, например, нервная, и верхние слои кожи. Внутренний слой (энтодерма) состоит из почти не пигментированных клеток, сохраняющих некоторое количество желтка; они дают начало главным образом тканям, выстилающим пищеварительный тракт и его производные. Следует, однако, подчеркнуть, что глубоких различий между этими двумя зародышевыми листками не существует. Эктодерма дает начало энтодерме, и если у некоторых форм границу между ними в области губы бластопора можно определить, то у других она практически неразличима. В экспериментах по трансплантации было показано, что различие между этими тканями определяется только их местоположением. Если участки, которые в норме оставались бы эктодермальными и дали бы начало производным кожи, пересадить на губу бластопора, они вворачиваются внутрь и становятся энтодермой, которая может превратиться в выстилку пищеварительного тракта, легкие или щитовидную железу.
Часто с появлением первичной кишки центр тяжести зародыша смещается, он начинает поворачиваться в своих оболочках, и в нем впервые устанавливаются передне-задняя (голова – хвост) и дорсо-вентральная (спина – живот) оси симметрии будущего организма.
Дробление негабаритов
Часто в рабочее пространство щековых дробилок попадают крупные куски округлой формы или куски, размер которых превышает ширину загрузочного отверстия. Крепкий кусок округлой формы не захватывается щеками и не дробится. Подвижная щека скользит по его поверхности и при приближении к неподвижной выталкивает его вверх. При отходе щеки кусок под тяжестью своей массы снова опускается вниз. Кусок большого размера может даже перекрыть загрузочное отверстие. В том и другом случае необходимо или принять меры, обеспечивающие прохождение куска через дробилку, т.е. раздробить его, или удалить этот кусок. Для этих целей изготавливают специальные устройства и инструменты:
- клин;
- «кошки»;
- клещи и т.п.
При дроблении крепкого куска округлой формы при помощи клина, клин, подвешенный на длинный трос на крюк крана, опускается в щель между куском и неподвижной или подвижной щекой. В результате изменяется угол захвата куска между щеками, и кусок может быть раздроблен. Если использование клина не дает желаемых результатов или загрузочное отверстие дробилки перекрыто куском, имеющим размер больший размера загрузочного отверстия, то кусок поворачивают при помощи так называемых «кошек» или специально для этой цели изготовленных клещей. «Кошки» в виде крюка большого размера с зубьями или без них вырезают обычно из листа железа толщиной 30-50 мм и подвешивают на крюк крана при помощи троса. В том случае, когда раздробить кусок не удается, его удаляют из рабочего пространства, обвязав тросом, концы которого закрепляются на крюке крана.
Все работы по дроблению или удалению крупного куска могут производиться при работающей дробилке, но с обязательным соблюдением правил техники безопасности, предусмотренных в специально разработанной для выполнения этих работ инструкции. Поэтому в здании, где работает щековая дробилка, всегда должны быть в наличии исправный, готовый к работе кран и указанные приспособления.
В случае забивки рабочего пространства щековой дробилки мелким материалом очистка производится при остановленной дробилке с выполнением правил техники безопасности. Очистку производят сверху. Материал вручную нагружают в бадью с открывающимся днищем, которую при помощи крана поднимают из рабочего пространства и выгружают.
Очистка рабочего пространства конусной дробилки для среднего и мелкого дробления производится при помощи крючков и пик, специально изготовленных для этой цели. При спуске в рабочее пространство дробилки необходимо пользоваться предохранительными поясами. Загрузочные отверстия в этом случае перекрываются настилом для предотвращения падения на людей случайных предметов. Разбутовка, очистка и пуск дробилок после их аварийной остановки должны производиться по специальной инструкции, утвержденной главным инженером фабрики, а резка металла, застрявшего в рабочей зоне, – только под наблюдением лица технического надзора. Иногда удаление застрявших в рабочем пространстве дробилки кусков руды осуществляют с помощью взрыва, при этом необходимо соблюдать Правила безопасности при взрывных работах. Разбивать в рабочем пространстве дробилки куски руды молотками или кувалдами запрещается.
Внезародышевые оболочки.
У животных, откладывающих яйца на суше или живородящих, зародышу необходимы дополнительные оболочки, защищающие его от обезвоживания (если яйца откладываются на суше) и обеспечивающие питание, удаление конечных продуктов обмена и газообмен.
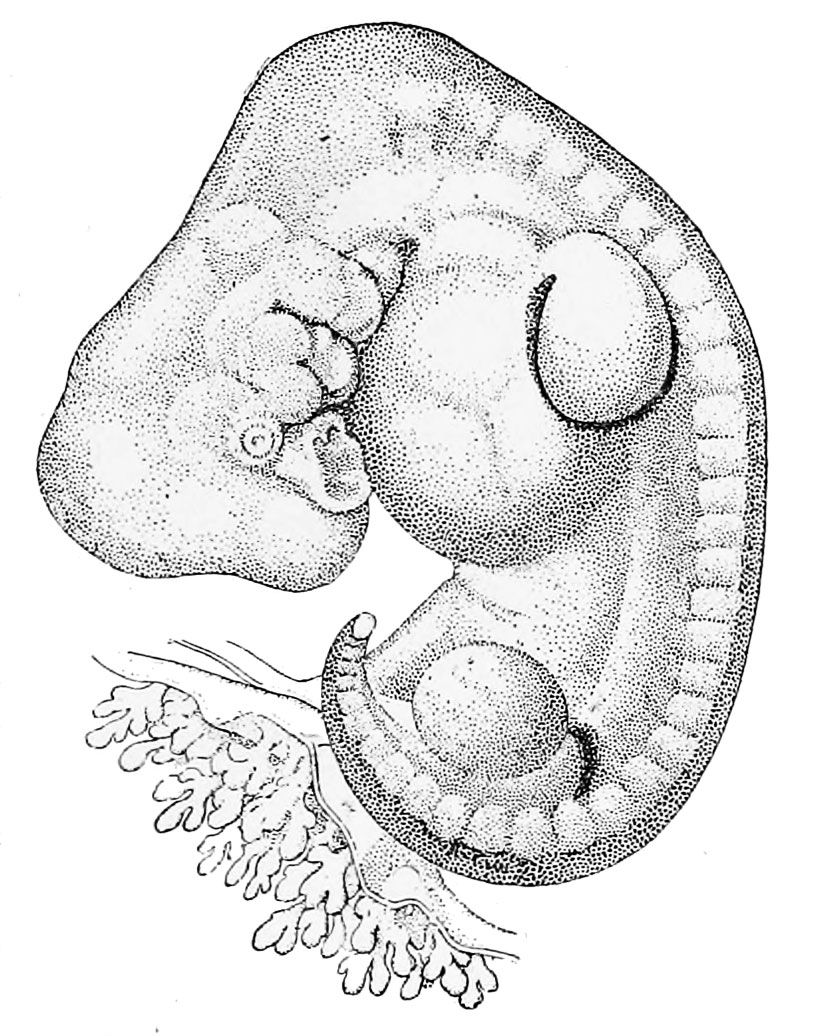
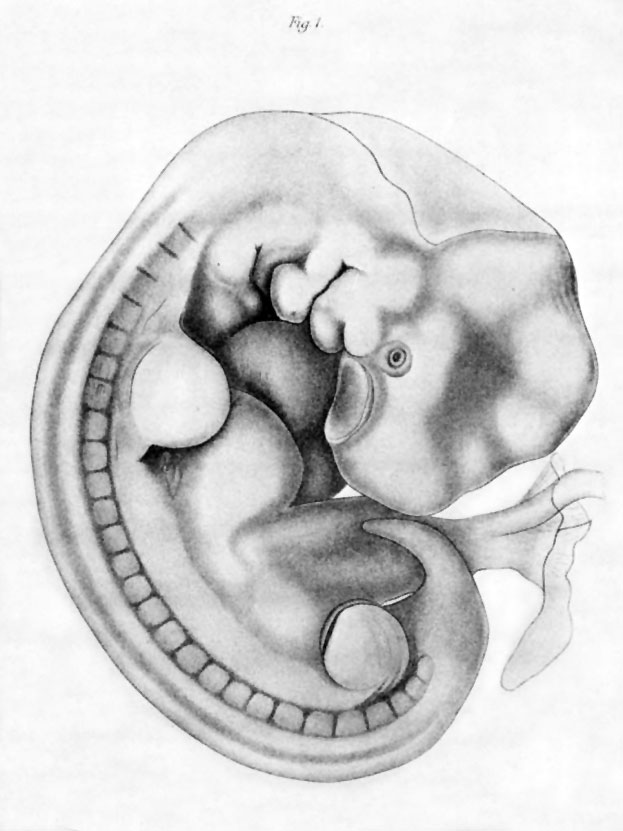
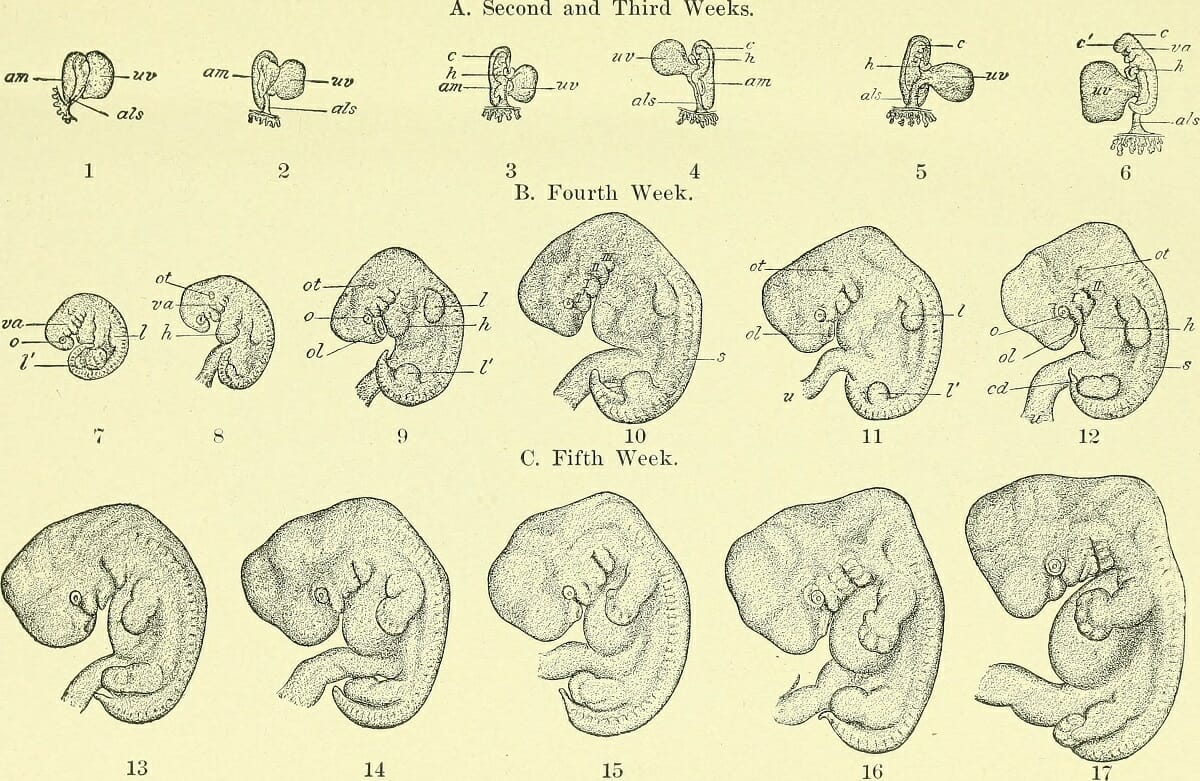
Эти функции выполняют внезародышевые оболочки – амнион, хорион, желточный мешок и аллантоис, образующиеся в процессе развития у всех пресмыкающихся, птиц и млекопитающих. Хорион и амнион тесно связаны между собой по происхождению; они развиваются из соматической мезодермы и эктодермы. Хорион – самая наружная оболочка, окружающая зародыш и три другие оболочки; эта оболочка проницаема для газов и через нее происходит газообмен. Амнион предохраняет клетки зародыша от высыхания благодаря амниотической жидкости, секретируемой его клетками. Желточный мешок, наполненный желтком, вместе с желточным стебельком поставляет зародышу подвергшиеся перевариванию питательные вещества; эта оболочка содержит густую сеть кровеносных сосудов и клетки, вырабатывающие пищеварительные ферменты. Желточный мешок, как и аллантоис, образуется из спланхнической мезодермы и энтодермы: энтодерма и мезодерма распространяются по всей поверхности желтка, обрастая его, так что в конце концов весь желток оказывается в желточном мешке. У пресмыкающихся и птиц аллантоис служит резервуаром для конечных продуктов обмена, поступающих из почек зародыша, а также обеспечивает газообмен. У млекопитающих эти важные функции выполняет плацента – сложный орган, образуемый ворсинками хориона, которые, разрастаясь, входят в углубления (крипты) слизистой оболочки матки, где вступают в тесный контакт с ее кровеносными сосудами и железами.
У человека плацента полностью обеспечивает дыхание зародыша, питание и выделение продуктов обмена в кровоток матери.
Внезародышевые оболочки не сохраняются в постэмбриональном периоде. У пресмыкающихся и птиц при вылуплении высохшие оболочки остаются в скорлупе яйца. У млекопитающих плацента и остальные внезародышевые оболочки выбрасываются из матки (отторгаются) после рождения плода. Эти оболочки обеспечили высшим позвоночным независимость от водной среды и, несомненно, сыграли важную роль в эволюции позвоночных, особенно в возникновении млекопитающих.
Правила дробления.
Установлено, что дробление подчиняется определенным правилам, названным именами исследователей, которые их впервые сформулировали. Правило Пфлюгера: веретено всегда тянется в направлении наименьшего сопротивления. Правило Бальфура: скорость голобластического дробления обратно пропорциональна количеству желтка (желток затрудняет деление как ядра, так и цитоплазмы). Правило Сакса: клетки обычно делятся на равные части, и плоскость каждого нового деления пересекает плоскость предшествующего деления под прямым углом. Правило Гертвига: ядро и веретено обычно располагаются в центре активной протоплазмы. Ось каждого веретена деления располагается по длинной оси массы протоплазмы. Плоскости деления обычно пересекают массу протоплазмы под прямым углом к ее осям.
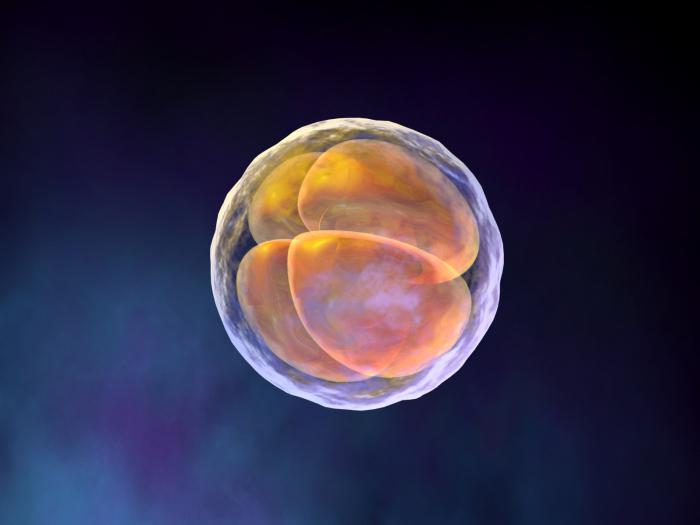

В результате дробления оплодотворенных яиц любого типа образуются клетки, называемые бластомерами. Когда бластомеров становится много (у земноводных, например, от 16 до 64 клеток), они образуют структуру, напоминающую ягоду малины и названную морулой.

Метамерия (расчленение тела на сходные сегменты).
Наибольшего успеха в длительном процессе эволюции достигли животные с сегментированным телом. Метамерное строение кольчатых червей и членистоногих отчетливо видно на протяжении всей их жизни. У большинства позвоночных первоначально сегментированное строение в дальнейшем становится мало различимым, однако на эмбриональных стадиях метамерия у них ясно выражена.
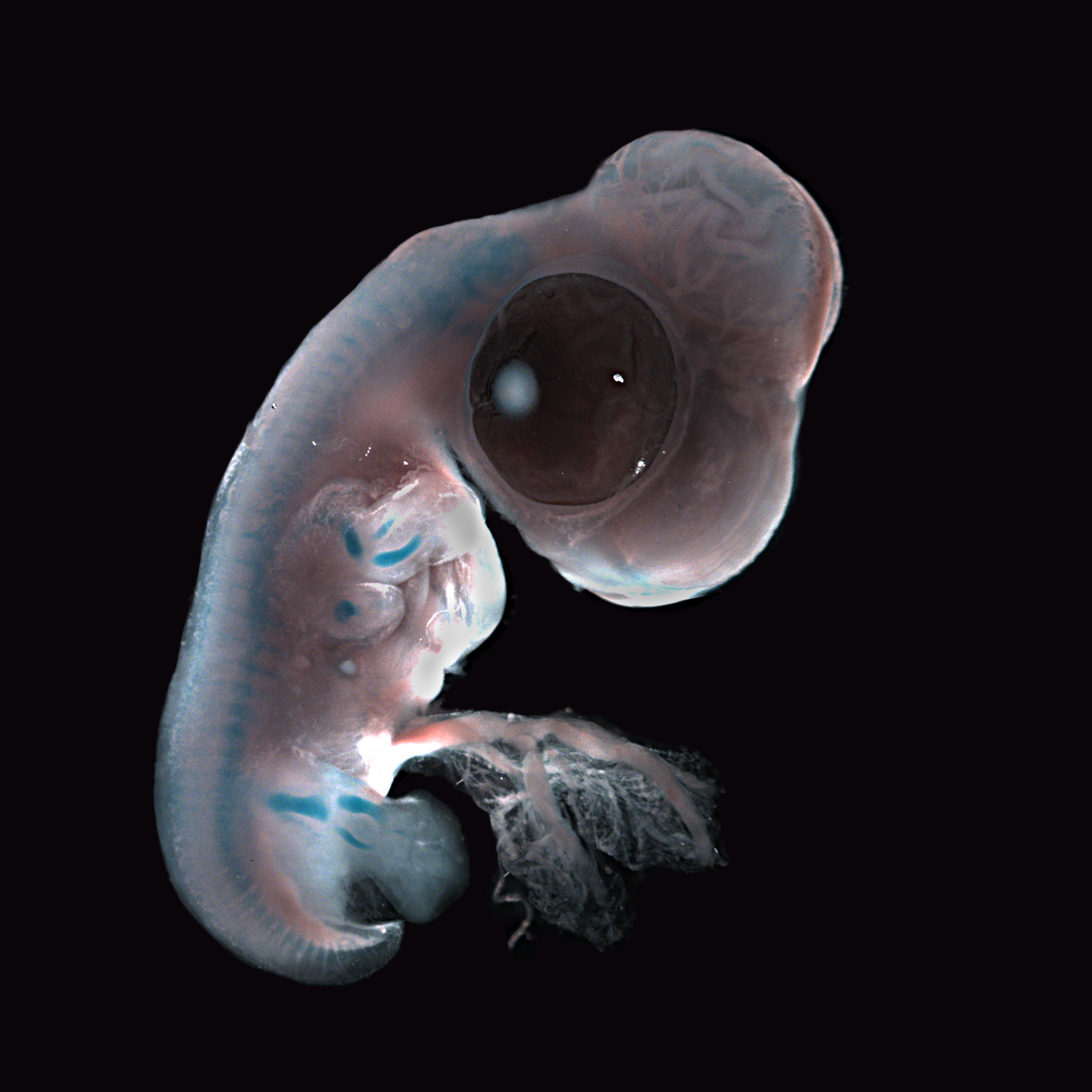
У ланцетника метамерия проявляется в строении целома, мышц и гонад. Для позвоночных характерно сегментарное расположение некоторых частей нервной, выделительной, сосудистой и опорной систем; однако уже на ранних стадиях эмбрионального развития на эту метамерию накладывается опережающее развитие переднего конца тела – т.н. цефализация. Если рассмотреть выращенного в инкубаторе 48-часового зародыша цыпленка, то можно выявить у него одновременно и двустороннюю симметрию и метамерию, наиболее отчетливо выраженную на переднем конце тела. Например, группы мышц, или сомиты, сначала появляются в области головы и образуются последовательно, так что наименее развитыми сегментированными сомитами оказываются задние.
Характерные черты дробления
Дробление как особый этап онтогенеза животных имеет характерные черты, которые свойственны большинству животных, но могут отсутствовать у некоторых групп.
- Бластомерs делятся очень быстро (у дрозофилы — раз в 20 минут) и более или менее синхронно.
- Интерфаза сокращена до S-фазы; в связи с этим транскрипция собственных генов зародыша полностью подавлена, транскрибируются только запасённые в яйцеклетке материнские мРНК.
- Между делениями нет периода роста, так что общая масса зародыша не растёт.
По всем этим характеристикам дробление млекопитающих резко отклоняется от типичного. Бластомеры делятся у них медленно, синхронность нарушается уже после 1—2 делений, в это же время активируется собственный геном зародыша.
Классификация типов дробления
На основе ряда существенных характеристик (степень детерминированности, полнота, равномерность и симметрия деления) выделяют ряд типов дробления. Типы дробления во многом определяются распределением веществ (в том числе, желтка) по цитоплазме яйца и характером межклеточных контактов, которые устанавливаются между бластомерами.
Дробление может быть: детерминированным и регулятивным; полным (голобластическим) или неполным (меробластическим); равномерным (бластомеры более-менее одинаковы по величине) и неравномерным (бластомеры не одинаковы по величине, выделяются две — три размерные группы, обычно называемые макро- и микромерами); наконец, по характеру симметрии различают радиальное, спиральное, различные варианты билатеризованных и анархическое дробление. В каждом из этих типов выделяют ряд вариантов.
Голобластическое дробление
Плоскости дробления разделяют яйцо полностью. Выделяют полное равномерное дробление, при котором бластомеры не различаются по размерам (такой тип дробления характерен для гомолецитальных и алецитальных яиц), и полное неравномерное дробление, при котором бластомеры могут существенно различаться по размерам.
Такой тип дробления характерен для умеренно телолецитальных яиц.
Меробластическое дробление
Дискоидальное (олигоцентрическое, потиентидальное, полиморфогистальное, винилдистерилориальное, компромертарионное)
- ограничено относительно небольшим участком у анимального полюса,
- плоскости дробления не проходят через всё яйцо и не захватывают желток.
Такой тип дробления типичен для телолецитальных яиц, богатых желтком (птицы, рептилии). Такое дробление называют также дискоидальным, так как в результате дробления на анимальном полюсе образуется небольшой диск клеток (бластодиск).
Поверхностное
- ядро зиготы делится в центральном островке цитоплазмы,
- получающиеся ядра перемещаются на поверхность яйца, образуя поверхностный слой ядер (синцитиальную бластодерму) вокруг лежащего в центре желтка. Затем ядра разделяются мембранами, и бластодерма становится клеточной.
Такой тип дробления наблюдается у членистоногих.
По типу симметрии дробящегося яйца
Радиальное
Ось яйца является осью радиальной симметрии. Типично для ланцетника, осетровых, амфибий, иглокожих, круглоротых.
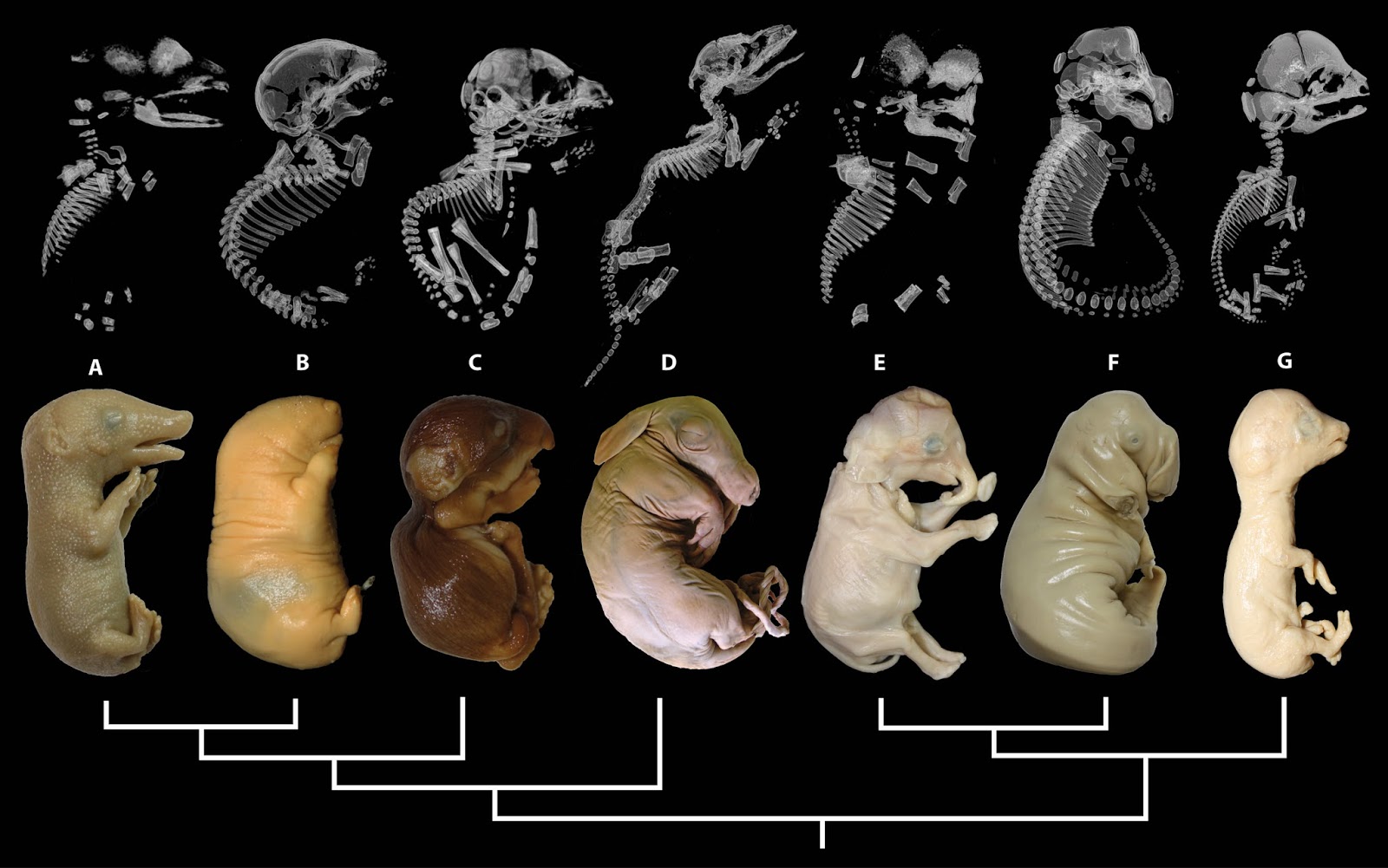


Хотя дробление амфибий часто описывается как радиальное, на самом деле еще до дробления, сразу же после оплодотворения, их яйцо приобретает билатеральную симметрию — на будущей спинной стороне зародыша образуется «серый серп» (его материал позднее попадает в клетки шпемановского организатора, или дорзальной губы бластопора). Билатеральную симметрию имеет и бластула амфибий.
Спиральное
В анафазе бластомеры разворачиваются. Отличается лево-правой дисимметрией (энантиоморфизм) уже на стадии четырёх (иногда двух) бластомеров. Типично для некоторых моллюсков, кольчатых и ресничных червей.
Анархическое
Бластомеры слабо связаны между собой, сначала образуют цепочки или бесформенную массу; часто у одного вида встречаются разные варианты расположения бластомеров. Типично для кишечнополостных.
| Это заготовка статьи по эмбриологии. Вы можете помочь проекту, дополнив её. |
| Это заготовка статьи по биологии. Вы можете помочь проекту, дополнив её.Это примечание по возможности следует заменить более точным. |
Зародышевые листки.
Эктодерму, энтодерму и мезодерму различают на основании двух критериев. Во-первых, по их местоположению в зародыше на ранних стадиях его развития: в этот период эктодерма всегда расположена снаружи, энтодерма – внутри, а мезодерма, появляющаяся последней, – между ними. Во-вторых, по их будущей роли: каждый из этих листков дает начало определенным органам и тканям, и их нередко идентифицируют по их дальнейшей судьбе в процессе развития. Однако напомним, что в период возникновения этих листков никаких принципиальных различий между ними не существует. В опытах по пересадке зародышевых листков было показано, что первоначально каждый из них обладает потенциями любого из двух других. Таким образом, их разграничение искусственно, но им очень удобно пользоваться при изучении эмбрионального развития.
Мезодерма, т.е. средний зародышевый листок, образуется несколькими способами. Она может возникать непосредственно из энтодермы путем образования целомических мешков, как у ланцетника; одновременно с энтодермой, как у лягушки; или путем деламинации, из эктодермы, как у некоторых млекопитающих. В любом случае вначале мезодерма представляет собой слой клеток, лежащих в пространстве, которое первоначально занимал бластоцель, т.е. между эктодермой с наружной и энтодермой с внутренней стороны.
Мезодерма вскоре расщепляется на два клеточных слоя, между которыми образуется полость, называемая целомом. Из этой полости в последующем образуются полость перикарда, окружающая сердце, плевральная полость, окружающая легкие, и брюшная полость, в которой лежат органы пищеварения. Наружный слой мезодермы – соматическая мезодерма – образует вместе с эктодермой т.н. соматоплевру. Из наружной мезодермы развиваются поперечнополосатые мышцы туловища и конечностей, соединительная ткань и сосудистые элементы кожи. Внутренний слой мезодермальных клеток называется спланхнической мезодермой и вместе с энтодермой образует спланхноплевру. Из этого слоя мезодермы развиваются гладкие мышцы и сосудистые элементы пищеварительного тракта и его производных. В развивающемся зародыше много рыхлой мезенхимы (эмбриональной мезодермы), заполняющей пространство между эктодермой и энтодермой.
У хордовых в процессе развития образуется продольный столбик плоских клеток – хорда, основной отличительный признак этого типа. Клетки хорды происходят из эктодермы у одних животных, из энтодермы у других и из мезодермы у третьих. В любом случае эти клетки уже на очень ранней стадии развития можно отличить от остальных, и расположены они в виде продольного столбика над первичной кишкой. У зародышей позвоночных хорда служит центральной осью, вокруг которой развивается осевой скелет, а над ней – центральная нервная система. У большинства хордовых это чисто эмбриональная структура, и только у ланцетника, круглоротых и пластиножаберных она сохраняется в течение всей жизни. Почти у всех других позвоночных клетки хорды замещаются костными клетками, образующими тело развивающихся позвонков; из этого следует, что наличие хорды облегчает формирование позвоночного столба.
Телолецитальные яйца
(от греч. télos – конец) содержат значительное количество желтка, а цитоплазма сконцентрирована у них на одном конце, обозначаемом обычно как анимальный полюс. Противоположный полюс, на котором сконцентрирован желток, называют вегетативным. Такие яйца типичны для кольчатых червей, головоногих моллюсков, бесчерепных (ланцетник), рыб, земноводных, пресмыкающихся, птиц и однопроходных млекопитающих. У них хорошо выражена анимально-вегетативная ось, определяемая градиентом распределения желтка; ядро обычно располагается эксцентрически; в яйцах, содержащих пигмент, он также распределяется по градиенту, но, в отличие от желтка, его больше на анимальном полюсе.
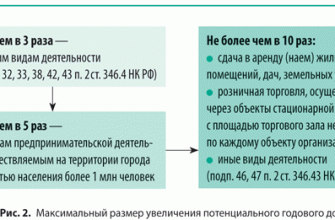
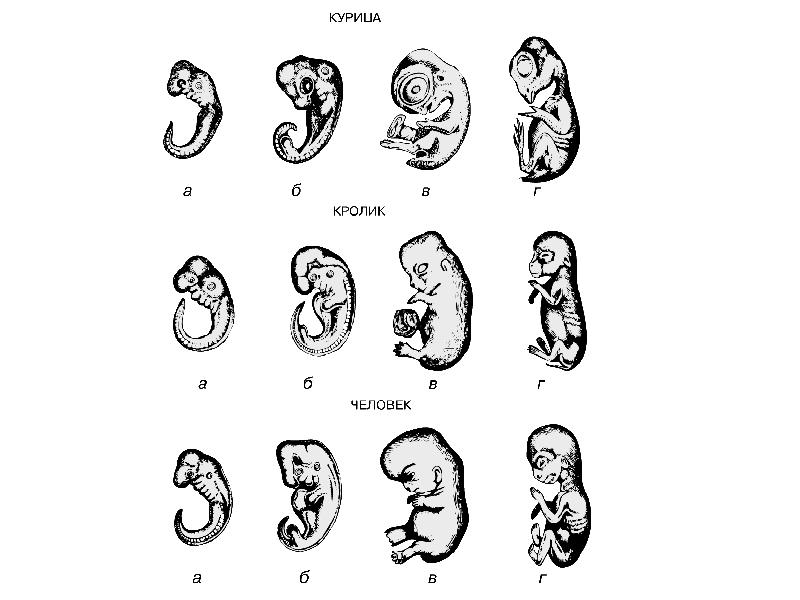
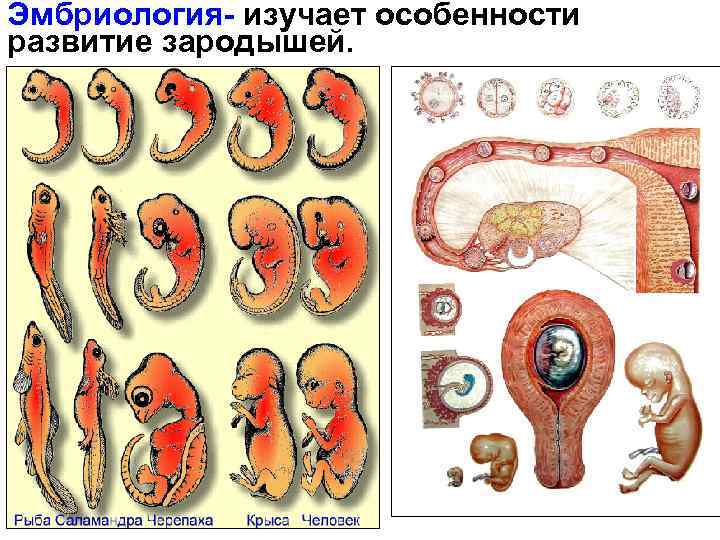
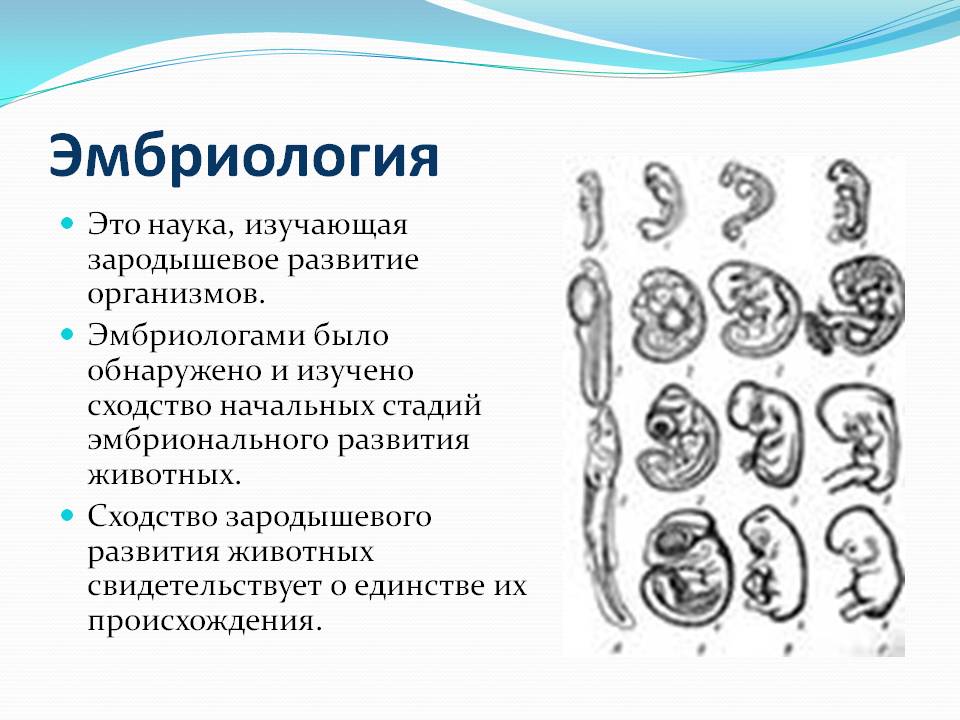
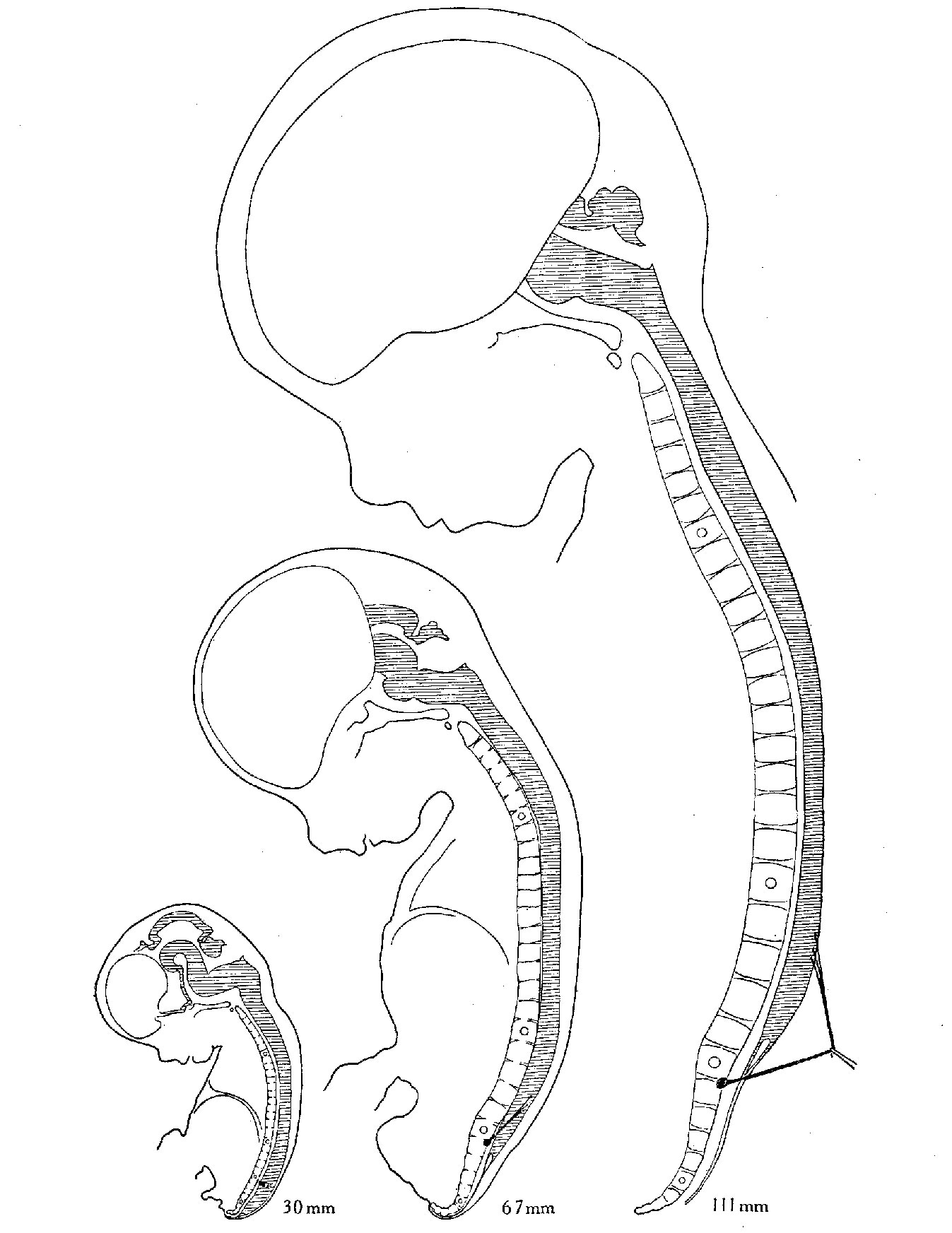
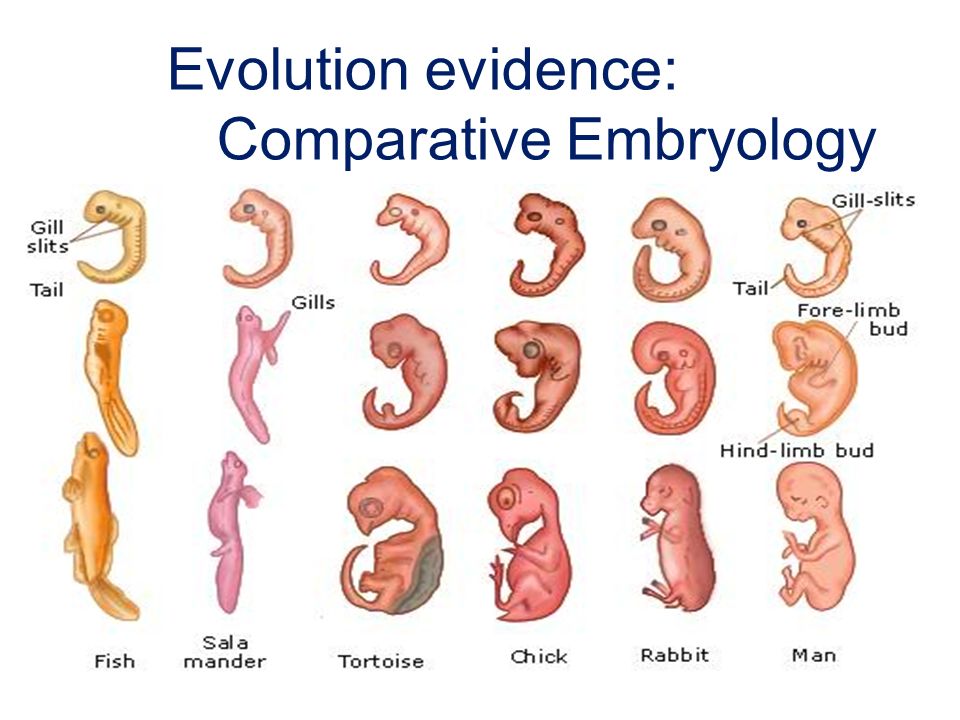
БИОГЕНЕТИЧЕСКИЙ ЗАКОН
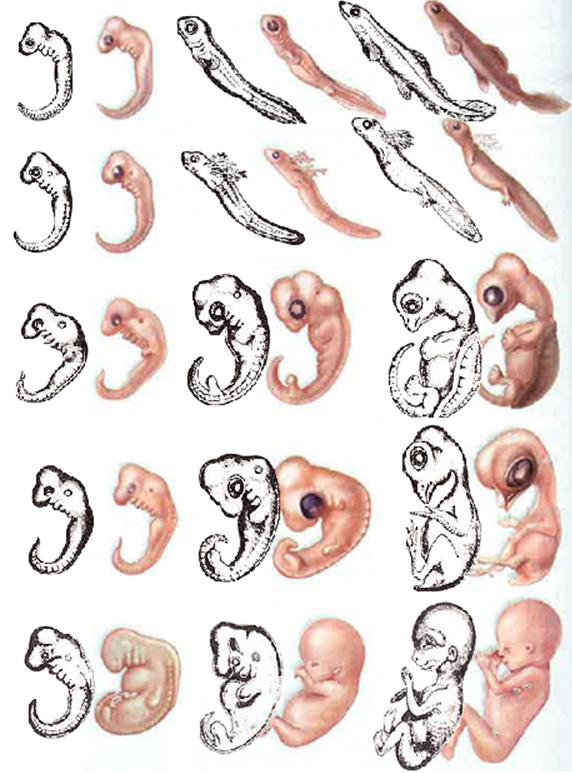
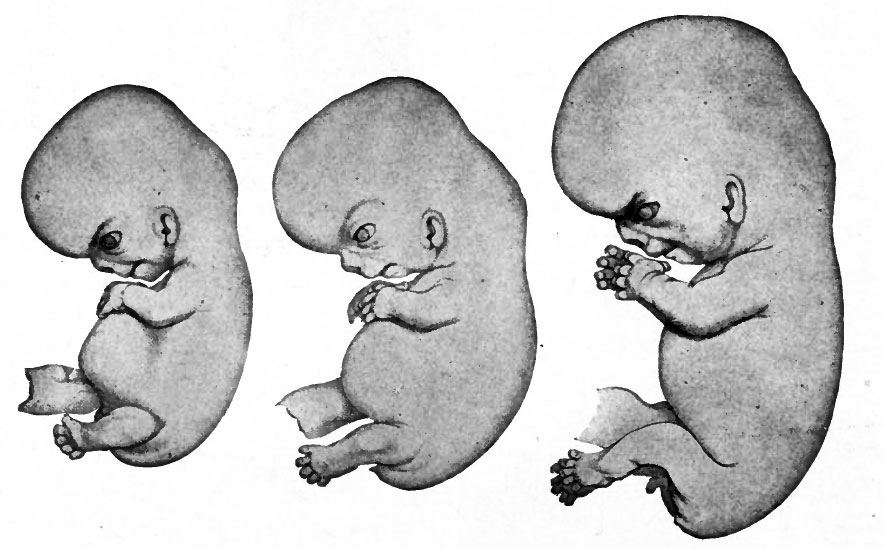
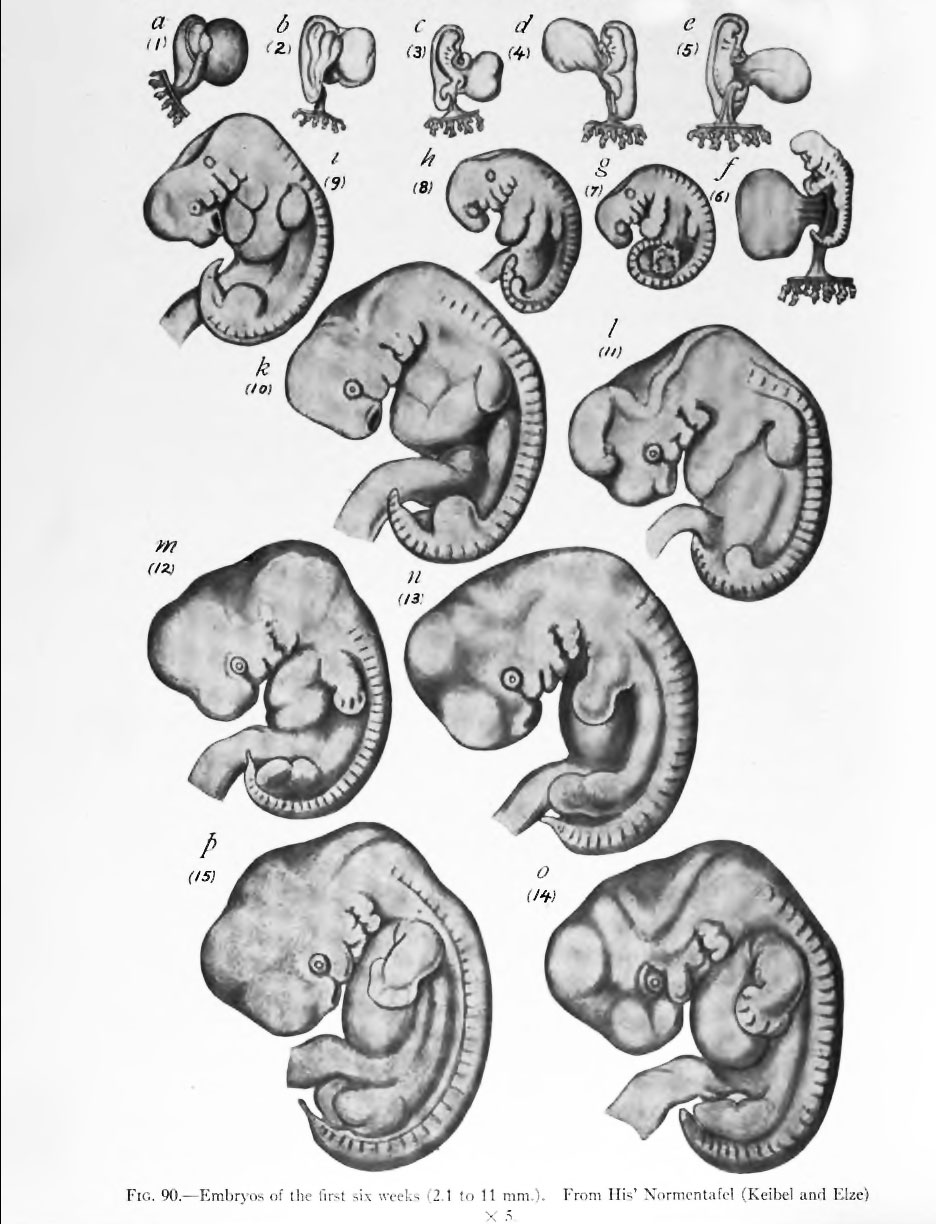
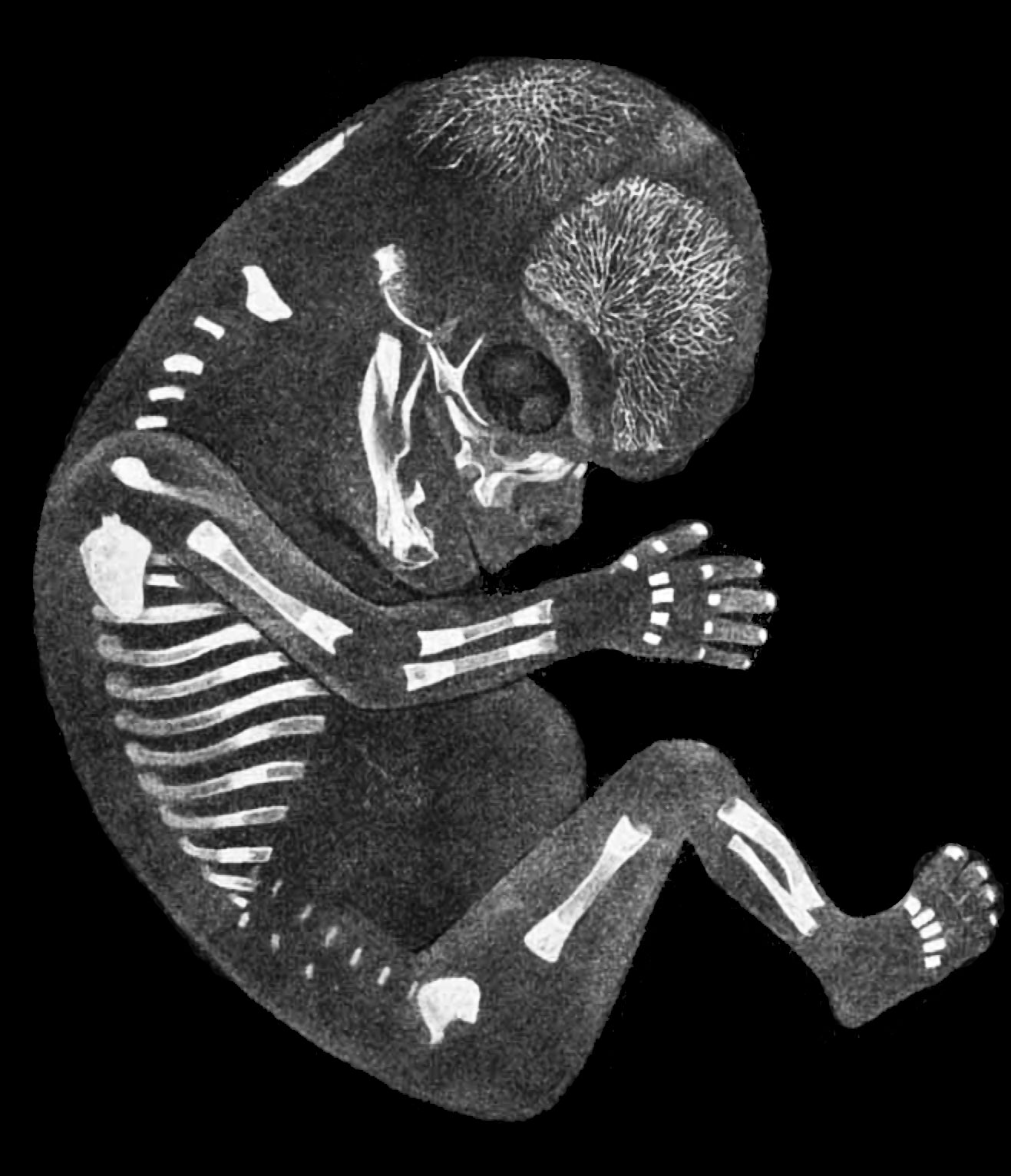
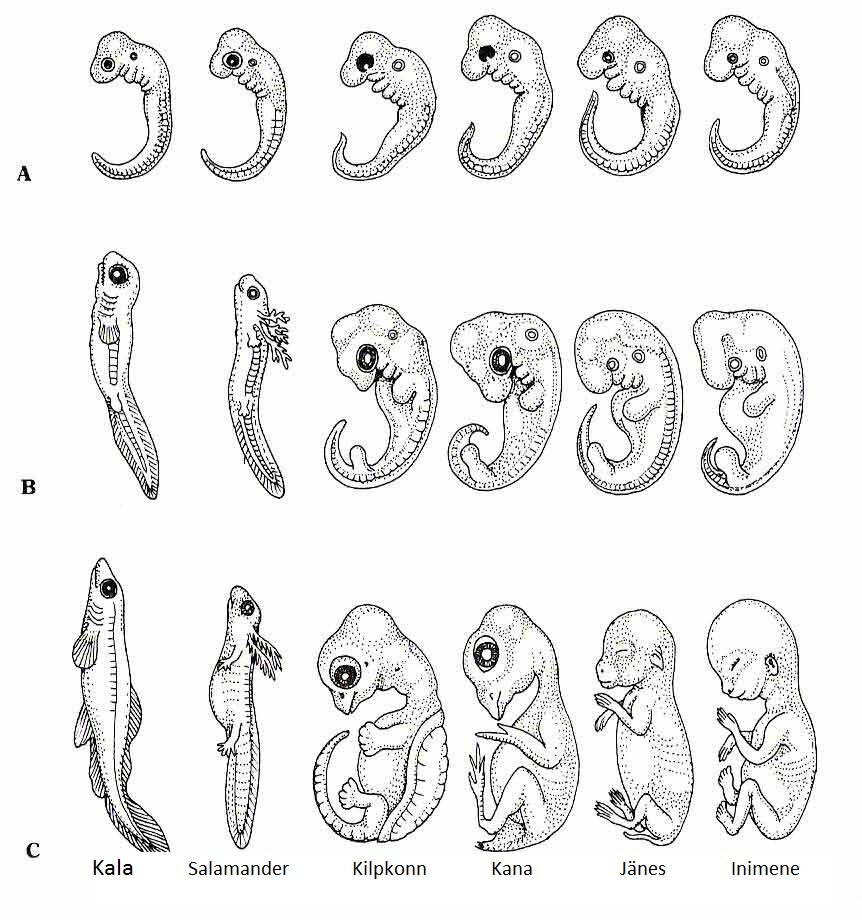
В 1828 К.фон Бэр сформулировал следующие положения: 1) наиболее общие признаки любой крупной группы животных появляются у зародыша раньше, чем менее общие признаки; 2) после формирования самых общих признаков появляются менее общие и так до появления особых признаков, свойственных данной группе; 3) зародыш любого вида животных по мере развития становится все менее похожим на зародышей других видов и не проходит через поздние стадии их развития; 4) зародыш высокоорганизованного вида может обладать сходством с зародышем более примитивного вида, но никогда не бывает похож на взрослую форму этого вида.
Биогенетический закон, сформулированный в этих четырех положениях, часто истолковывают неверно. Закон этот просто утверждает, что некоторые стадии развития высокоорганизованных форм обладают явным сходством с некоторыми стадиями развития нижестоящих на эволюционной лестнице форм. Предполагается, что это сходство можно объяснить происхождением от общего предка. О взрослых стадиях низших форм ничего не говорится. В данной статье сходство между зародышевыми стадиями подразумевается; в противном случае развитие каждого вида пришлось бы описывать отдельно.
По-видимому, в длительной истории жизни на Земле среда играла главную роль в отборе зародышей и взрослых организмов, наиболее приспособленных для выживания. Узкие рамки, создаваемые средой в отношении возможных колебаний температуры, влажности и снабжения кислородом, сокращали разнообразие форм, приводя их к относительно общему типу. В результате возникло то сходство строения, которое лежит в основе биогенетического закона, если речь идет о зародышевых стадиях. Разумеется, у ныне существующих форм в процессе зародышевого развития проявляются особенности, соответствующие времени, месту и способам размножения данного вида.
Онтогенез, т.е. развитие отдельной особи, предваряет филогенез, т.е. развитие группы, потому что мутации обычно возникают в половых клетках до оплодотворения. Изменения в эмбрионе естественно предшествуют изменениям взрослой особи, имеющим эволюционное значение, а часто и вызывают их. Новая особь «закладывается» в момент оплодотворения, а зародышевое развитие только подготавливает его к превратностям взрослого существования и созданию будущих зародышей. См. также ЦИТОЛОГИЯ; НАСЛЕДСТВЕННОСТЬ; СИСТЕМАТИКА ЖИВОТНЫХ.



Бластула.
По мере продолжения дробления бластомеры становятся все мельче и все плотнее прилегают друг к другу, приобретая гексагональную форму. Такая форма повышает структурную жесткость клеток и плотность слоя. Продолжая делиться, клетки раздвигают друг друга и в итоге, когда их число достигает нескольких сотен или тысяч, формируют замкнутую полость – бластоцель, в который поступает жидкость из окружающих клеток. В целом это образование носит название бластулы. Ее формированием (в котором клеточные движения не участвуют) завершается период дробления яйца.
В гомолецитальных яйцах бластоцель может располагаться в центре, но в телолецитальных яйцах он обычно бывает сдвинут желтком и располагается эксцентрически, ближе к анимальному полюсу и прямо под бластодиском. Итак, бластула обычно представляет собой полый шарик, полость которого (бластоцель) заполнена жидкостью, но в телолецитальных яйцах с дискоидальным дроблением бластула представлена уплощенной структурой.
При голобластическом дроблении стадия бластулы считается завершенной, когда в результате деления клеток соотношение между объемами их цитоплазмы и ядра становится таким же, как в соматических клетках. В оплодотворенном яйце объемы желтка и цитоплазмы совершенно не соответствуют размерам ядра. Однако в процессе дробления количество ядерного материала несколько увеличивается, тогда как цитоплазма и желток только делятся. В некоторых яйцах отношение объема ядра к объему цитоплазмы в момент оплодотворения составляет примерно 1:400, а к концу стадии бластулы – примерно 1:7. Последнее близко к соотношению, характерному и для первичной половой и для соматической клетки.
Поверхности поздней бластулы оболочников и земноводных можно картировать; для этого на разные ее участки наносят прижизненные (не наносящие вреда клеткам) красители – сделанные цветные метки сохраняются в ходе дальнейшего развития и позволяют установить, какие органы возникают из каждого участка. Эти участки называют презумптивными, т.е. такими, судьбу которых при нормальных условиях развития можно предсказать. Если, однако, на стадии поздней бластулы или ранней гаструлы переместить эти участки или поменять местами, их судьба изменится. Подобные эксперименты показывают, что до какой-то определенной стадии развития каждый бластомер способен превратиться в любую из множества разнообразных клеток, составляющих организм.

Голобластическое дробление
характерно для гомолецитальных яиц. Плоскости дробления разделяют яйцо полностью. Они могут делить его на равные части, как у морской звезды или морского ежа, или же на неравные части, как у брюхоногого моллюска Crepidula. Дробление умеренно телолецитального яйца ланцетника происходит по голобластическому типу, однако неравномерность деления проявляется только после стадии четырех бластомеров. У некоторых клеток после этой стадии дробление становится крайне неравномерным; образующиеся при этом мелкие клетки называют микромерами, а крупные клетки, содержащие желток, – макромерами. У моллюсков плоскости дробления проходят таким образом, что начиная со стадии восьми клеток бластомеры располагаются по спирали; этот процесс регулируется ядром.
Инволюция.
У пресмыкающихся и птиц, телолецитальные яйца которых содержат большое количество желтка и дробятся меробластически, клетки бластулы на очень небольшом участке приподнимаются над желтком и затем начинают вворачиваться внутрь, под клетки верхнего слоя, образуя второй (нижний) слой. Этот процесс вворачивания клеточного пласта называют инволюцией. Верхний слой клеток становится наружным зародышевым листком, или эктодермой, а нижний – внутренним, или энтодермой. Эти слои переходят один в другой, а место, где происходит переход, известно под названием губы бластопора. Крыша первичной кишки у зародышей этих животных состоит из вполне сформировавшихся энтодермальных клеток, а дно – из желтка; дно из клеток образуется позднее.
Оплодотворение.
Оплодотворение – сложный процесс, в ходе которого сперматозоид проникает в яйцо и их ядра сливаются. В результате слияния гамет образуется зигота – по существу уже новая особь, способная развиваться при наличии необходимых для этого условий. Оплодотворение вызывает активацию яйца, стимулируя его к последовательным изменениям, приводящим к развитию сформированного организма. При оплодотворении происходит также амфимиксис, т.е. смешение наследственных факторов в результате слияния ядер яйца и сперматозоида. Яйцо обеспечивает половину необходимых хромосом и обычно все питательные вещества, необходимые для ранних стадий развития.
При соприкосновении сперматозоида с поверхностью яйца желточная оболочка яйца изменяется, превращаясь в оболочку оплодотворения. Это изменение считается доказательством того, что произошла активация яйца. Одновременно на поверхности яиц, содержащих мало желтка или не содержащих его вовсе, возникает т.н. кортикальная реакция, не допускающая проникновения в яйцо других сперматозоидов. У яиц, содержащих очень много желтка, кортикальная реакция возникает позднее, так что в них обычно проникает несколько сперматозоидов. Но даже в таких случаях оплодотворение совершает только один сперматозоид, первым дошедший до ядра яйца.
У некоторых яиц в месте соприкосновения сперматозоида с плазматической мембраной яйца образуется выпячивание мембраны – т.н. бугорок оплодотворения; он облегчает проникновение сперматозоида. Обычно в яйцо проникают головка сперматозоида и центриоли, находящиеся в его средней части, а хвост остается снаружи. Центриоли способствуют образованию веретена при первом делении оплодотворенного яйца. Процесс оплодотворения можно считать завершенным, когда два гаплоидных ядра – яйцеклетки и сперматозоида – сливаются и их хромосомы конъюгируют, готовясь к первому дроблению оплодотворенного яйца. См. также ЯЙЦО.
Классификация типов дробления
На основе ряда существенных характеристик (степень детерминированности, полнота, равномерность и симметрия деления) выделяют ряд типов дробления. Типы дробления во многом определяются распределением веществ (в том числе, желтка) по цитоплазме яйца и характером межклеточных контактов, которые устанавливаются между бластомерами.
Дробление может быть: детерминированным и регулятивным; полным (голобластическим) или неполным (меробластическим); равномерным (бластомеры более-менее одинаковы по величине) и неравномерным (бластомеры не одинаковы по величине, выделяются две — три размерные группы, обычно называемые макро- и микромерами); наконец, по характеру симметрии различают радиальное, спиральное, различные варианты билатеризованных и анархическое дробление. В каждом из этих типов выделяют ряд вариантов.


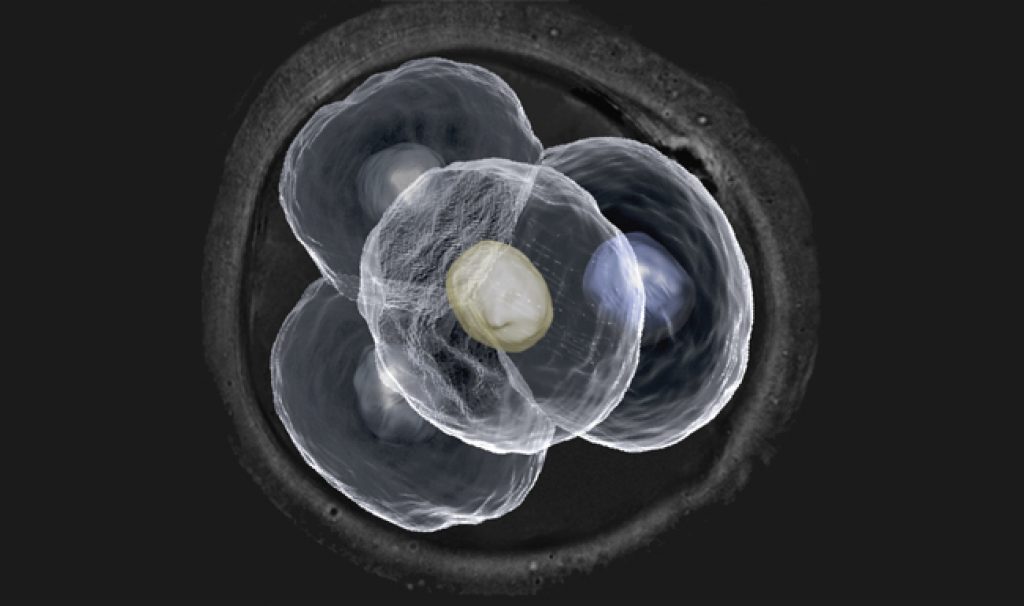
Голобластическое дробление
Плоскости дробления разделяют яйцо полностью. Выделяют полное равномерное дробление, при котором бластомеры не различаются по размерам (такой тип дробления характерен для гомолецитальных и алецитальных яиц), и полное неравномерное дробление, при котором бластомеры могут существенно различаться по размерам.
Такой тип дробления характерен для умеренно телолецитальных яиц.
Меробластическое дробление
Дискоидальное (олигоцентрическое, потиентидальное, полиморфогистальное, винилдистерилориальное, компромертарионное)
- ограничено относительно небольшим участком у анимального полюса,
- плоскости дробления не проходят через всё яйцо и не захватывают желток.
Такой тип дробления типичен для телолецитальных яиц, богатых желтком (птицы, рептилии). Такое дробление называют также дискоидальным, так как в результате дробления на анимальном полюсе образуется небольшой диск клеток (бластодиск).
Поверхностное
- ядро зиготы делится в центральном островке цитоплазмы,
- получающиеся ядра перемещаются на поверхность яйца, образуя поверхностный слой ядер (синцитиальную бластодерму) вокруг лежащего в центре желтка. Затем ядра разделяются мембранами, и бластодерма становится клеточной.
Такой тип дробления наблюдается у членистоногих.
По типу симметрии дробящегося яйца
Радиальное
Ось яйца является осью радиальной симметрии. Типично для ланцетника, осетровых, амфибий, иглокожих, круглоротых.
Хотя дробление амфибий часто описывается как радиальное, на самом деле еще до дробления, сразу же после оплодотворения, их яйцо приобретает билатеральную симметрию — на будущей спинной стороне зародыша образуется «серый серп» (его материал позднее попадает в клетки шпемановского организатора, или дорзальной губы бластопора). Билатеральную симметрию имеет и бластула амфибий.
Спиральное
В анафазе бластомеры разворачиваются. Отличается лево-правой дисимметрией (энантиоморфизм) уже на стадии четырёх (иногда двух) бластомеров. Типично для некоторых моллюсков, кольчатых и ресничных червей.
Анархическое
Бластомеры слабо связаны между собой, сначала образуют цепочки или бесформенную массу; часто у одного вида встречаются разные варианты расположения бластомеров. Типично для кишечнополостных.
Производные зародышевых листков.
Дальнейшая судьба трех зародышевых листков различна.
Из эктодермы развиваются: вся нервная ткань; наружные слои кожи и ее производные (волосы, ногти, зубная эмаль) и частично слизистая ротовой полости, полостей носа и анального отверстия.
Энтодерма дает начало выстилке всего пищеварительного тракта – от ротовой полости до анального отверстия – и всем ее производным, т.е. тимусу, щитовидной железе, паращитовидным железам, трахее, легким, печени и поджелудочной железе.
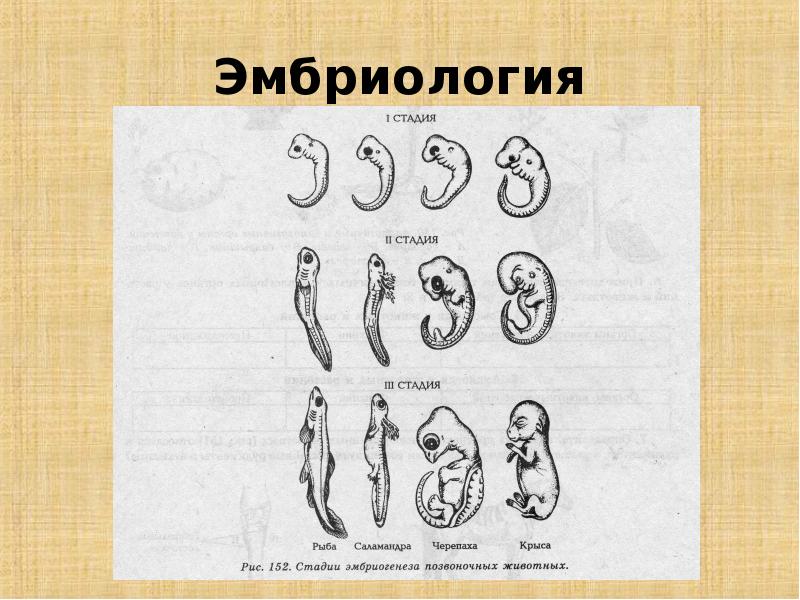


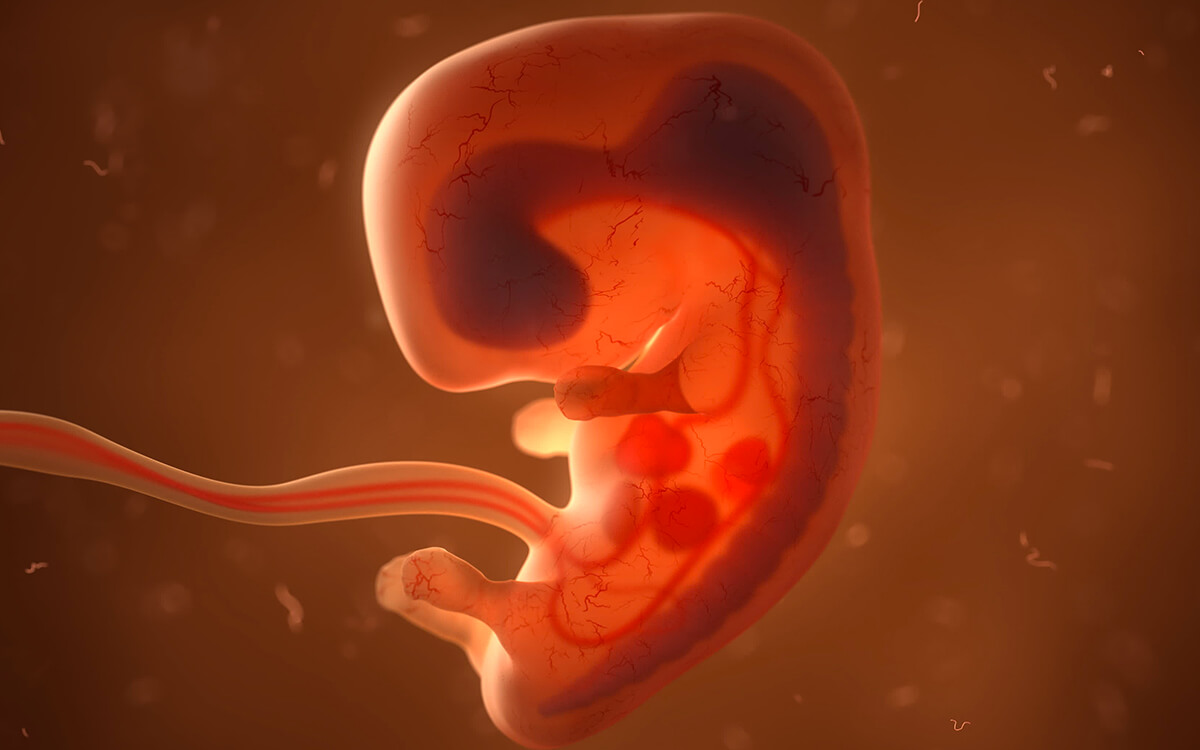




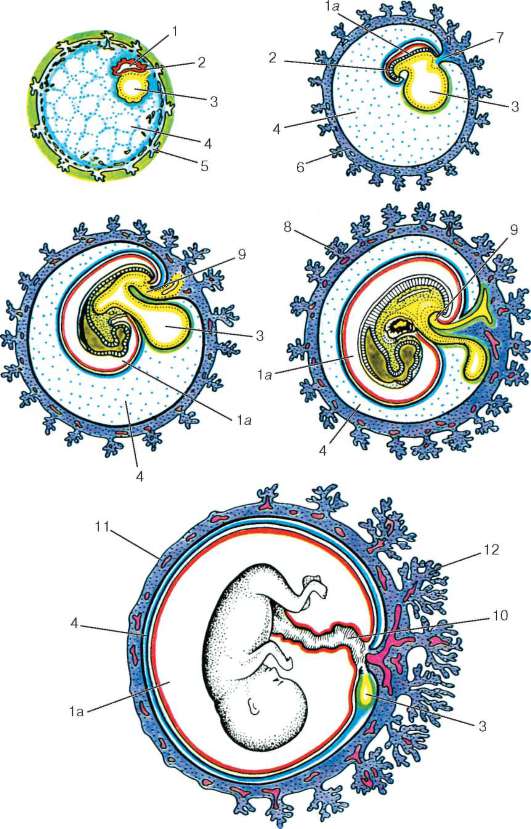

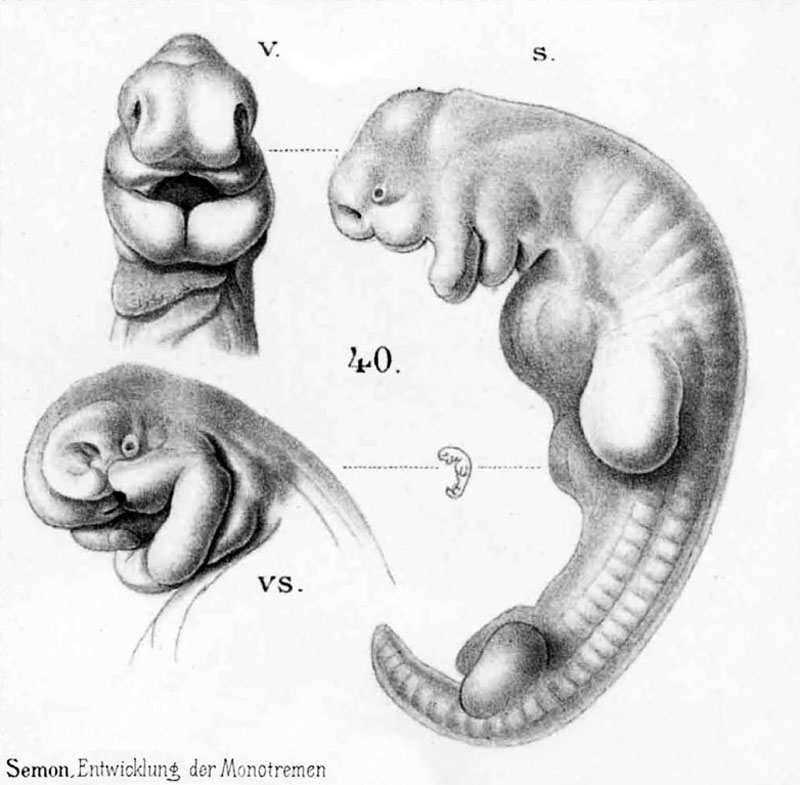
Из мезодермы образуются: все виды соединительной ткани, костная и хрящевая ткани, кровь и сосудистая система; все типы мышечной ткани; выделительная и репродуктивная системы, дермальный слой кожи.
У взрослого животного очень мало таких органов энтодермального происхождения, которые не содержали бы нервных клеток, происходящих из эктодермы
В каждом важном органе содержатся и производные мезодермы – кровеносные сосуды, кровь, часто и мышцы, так что структурная обособленность зародышевых листков сохраняется только на стадии их образования. Уже в самом начале своего развития все органы приобретают сложное строение, и в них входят производные всех зародышевых листков
Характерные черты дробления
Дробление как особый этап онтогенеза животных имеет характерные черты, которые свойственны большинству животных, но могут отсутствовать у некоторых групп.
- Бластомерs делятся очень быстро (у дрозофилы — раз в 20 минут) и более или менее синхронно.
- Интерфаза сокращена до S-фазы; в связи с этим транскрипция собственных генов зародыша полностью подавлена, транскрибируются только запасённые в яйцеклетке материнские мРНК.
- Между делениями нет периода роста, так что общая масса зародыша не растёт.
По всем этим характеристикам дробление млекопитающих резко отклоняется от типичного. Бластомеры делятся у них медленно, синхронность нарушается уже после 1—2 делений, в это же время активируется собственный геном зародыша.