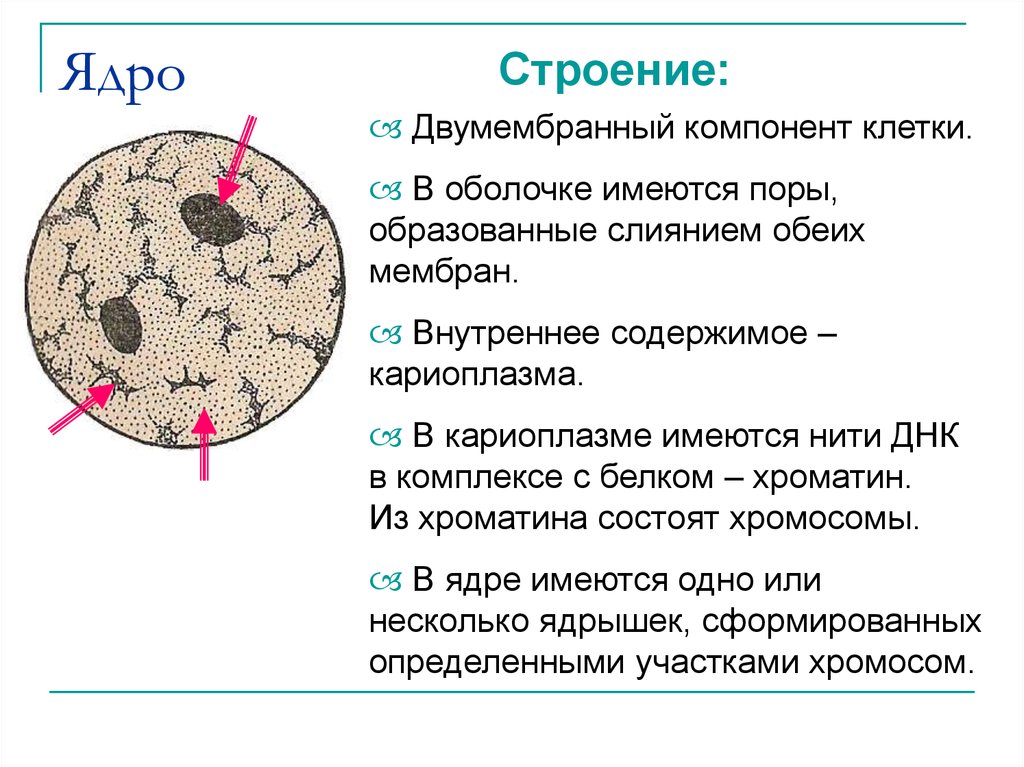
Половой хроматин в судебно-медицинском отношении
Исследование П. х. в суд.-мед. практике производится с целью установления половой принадлежности следов крови, слюны и других биол, жидкостей, вырванных волос, следов-отпечатков клеток тканей и органов, кусочков тканей, которые могут быть обнаружены на месте происшествия, на различных предметах, одежде, теле потерпевшего и подозреваемого в совершении преступления, на орудиях травмы, на транспортных средствах, а также при обнаружении обгоревших трупов или частей расчлененных трупов. Реже П. х. исследуют с целью суд.-мед. установления генетического пола у лиц с аномалиями полового развития, используя общепринятые методики.
Для приготовления препаратов из следов крови (см.) и слюны (см.) кусочки предмета-носителя помещают в пробирку и заливают 0,5—40% (следы крови) или 5—10% (следы слюны) уксусной к-той. Экстрагируют при комнатной температуре в течение нескольких часов и, удалив кусочки предмета-носителя, центрифугируют. Осадок переносят на предметное стекло и высушивают на воздухе. С пятен крови на предметах, не впитывающих жидкость (металл, стекло, пластмасса и др.), делают соскобы, которые затем обрабатывают таким же образом.
При исследовании волос (см.) корень волоса помещают на предметное стекло и добавляют 10—25% уксусную к-ту. После набухания отделяют и измельчают волосяной фолликул, удаляя остальные части волоса.
Из кусочков тканей и органов, при необходимости предварительно выдержав их до набухания в уксусной к-те соответствующей концентрации или в физиол, р-ре, готовят гистологические препараты, мазки или препараты-отпечатки. Следы-наложения клеток, тканей или органов на орудиях травмы смывают физиол, р-ром, одновременно соскабливая их. Мелкие кусочки тканей, встречающиеся в таких следах, измельчают препаровальными иглами. Смывы-соскобы помещают в пробирки, центрифугируют, из осадка готовят гистологические препараты. Исследование препаратов целесообразно начинать с выявления Y-хроматина, т. к. при его отсутствии те же препараты могут быть снова использованы для выявления X-хроматина. При исследовании учитывают только достаточно хорошо сохранившиеся неповрежденные ядра клеток. При анализе следов крови Y-хроматин определяют в ядрах лимфоцитов, т. к. в нейтрофилах Y-хроматин в препаратах из следов крови мужчин может не выявляться.
При отсутствии повышенной влажности Половой хроматин может длительно сохраняться в высохших следах, а также в клетках фолликула вырванного волоса. Высокая температура (выше 150°) разрушает ядра клеток и П. х. Значительная влажность в течение нескольких суток также приводит к разрушению клеток, что делает невозможным выявление полового хроматина. Т. к. условия, в к-рых находятся следы, могут последовательно меняться, решающее значение для установления пригодности следов крови, слюны и т. д. для определения П. х. имеет состояние обнаруживаемых в них клеток и их ядер. В клетках высохших кусочков тканей, не подвергающихся действию влаги, П. х. сохраняется длительное время. В целых трупах и в их крупных частях в процессе аутолиза и гниения в течение нескольких суток происходит деструкция клеточных ядер. В обгоревших трупах половой хроматин нек-рое время может сохраняться в клетках глубоко расположенных органов и тканей.
При выявлении небольшого числа клеток, сохранивших ядра, исследуемых на Половой хроматин, с целью установления статистической достоверности результатов используют различные математические методы анализа, учитывающие как общее число обнаруженных клеток, так и число клеток. содержащих X- или Y-хроматин.
Библиография: Давиденкова Е. Ф., Берлинская Д. К. и Тысячнюк С. Ф. Клинические синдромы при аномалиях половых хромосом, Л., 1973; Захаров А. Ф. Хромосомы человека, М., 1977; Капустин А. В. Судебно-медицинская диагностика пола по половым различиям в клетках, М., 1969; Лабораторные и специальные методы исследования в судебной медицине, под ред. В. И. Пашковой и В. В. Томилина, с. 157, М., 1975; Любинская С. И. и Антонова С. Н. Исследование Y-хроматина в следах крови, Суд.-мед. экспертиза, т. 18, № 3, с. 17, 1975; Основы цитогенетики человека, под ред. A. А. Прокофьевой-Бельговской, М., 1969; Methods in human cytogenetics, ed. by H. G. Schwarzacher a. U. Wolf, p. 207, B. а. о., 1974; The sex chromatin, ed. by K. L. Moore, Philadelphia — L., 1966.
Структурные и функциональные компоненты хроматина
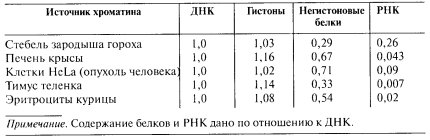
С целью определить, что такое хроматин на биохимическом уровне, ученые экстрагировали это вещество из клеток, переводили в раствор и в таком виде изучали компонентный состав и структуру. При этом использовались как химические, так и физические методы, включая технологии электронной микроскопии. Выяснилось, что химический состав хроматина на 40% представлен длинными молекулами ДНК и почти на 60% – различными белками. Последние подразделяются на две группы: гистоны и негистоновые.
Гистонами называют большое семейство основных ядерных белков, которые прочно связываются с ДНК, формируя структурный скелет хроматина. Их количество примерно равно процентному содержанию генетических молекул.
Остальная часть (до 20%) протеиновой фракции приходится на ДНК-связывающие и пространственно-модифицирующие белки, а также ферменты, принимающие участие в процессах считывания и копирования генетической информации.
Помимо основных элементов, в составе хроматина в небольшом количестве обнаруживаются рибонуклеиновые кислоты (РНК), гликопротеиды, углеводы и липиды, однако вопрос об их ассоциации с ДНК-упаковочным комплексом до сих пор открыт.
Роль вещества наследственности
Комплексы, включающие ДНК, РНК и белок, называемые хроматином, участвуют в онтогенезе клеток и изменяют свой состав в зависимости от типа ткани, а также от стадии развития организма в целом. Например, в эпителиальных клетках кожи такие гены, как эхансер и промотор, заблокированы белками-репрессорами, а эти же регуляторные гены в секреторных клетках эпителия кишечника активны и находятся в зоне открытого хроматина. Ученые-генетики установили, что на долю ДНК, не кодирующей белки, приходится более 95 % всего генома человека. Это значит, что управляющих генов намного больше, чем тех, которые ответственны за синтез пептидов. Внедрение таких методов, как ДНК-чипы и секвенирование, позволило выяснить, что такое хроматин, и, как следствие, провести картирование генома человека.
Исследования хроматина очень важны в таких отраслях науки, как генетика человека и медицинская генетика. Это связано с резко возросшим уровнем появления наследственных заболеваний — как генных, так и хромосомных. Раннее выявление этих синдромов повышает процент положительных прогнозов при их лечении.

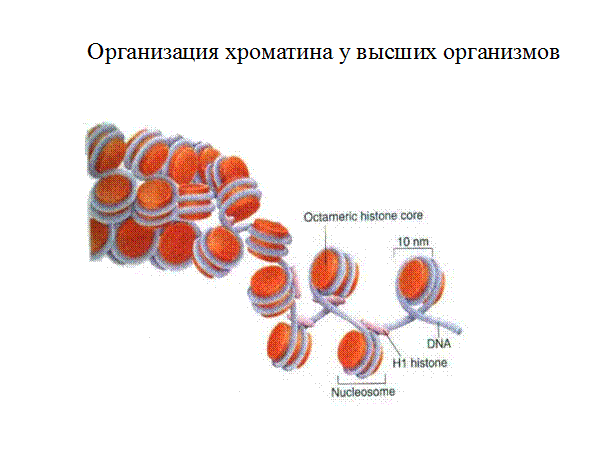





Значение открытого хроматина
В геномах эукариот некодирующие последовательности, участвующие в регуляции экспрессии генов на разных стадиях развития организма или в разных тканях, приобретают особое значение. Открытие и характеристика регуляторных участков становится необходимой для понимания закономерностей в экспрессии генов. Так в геноме человека более 95 % ДНК является некодирующей. В этот класс последовательностей, кроме мусорной ДНК, входят важные регуляторные последовательности: промоторы, энхансеры, сайленсеры, инсуляторы или (англ. locus control regions (LCR)). ENCODE консорциум показал, что сайты гиперчувствительности к ДНКазе I, идентифицированные в 1 % генома человека, являются маркерами гистонов, участков ранней репликации, сайтов начала транскрипции и транскрипционных факторов. Также открытый хроматин часто связан с активно транскрибируемыми некодирующими РНК.
Распределение открытого хроматина
Кроме некодирующих регуляторных последовательностей, открытый хроматин также ассоциирован с экзонами и интронами активно транскрибируемых генов. Особенно часто такие участки открытого хроматина совпадают первым экзоном и интроном гена. Однако наличие открытого хроматина не является достаточным условием активности гена. Нетранскрибируемые гены, связанные с открытым хроматином, находятся в состоянии «готовности» к транскрипции (англ. poised state). Таким образом, формирование открытого хроматина или перевод в неактивное состояние является важным для регуляции экспрессии генов.
Свободными от нуклеосом могут быть не только участки связывания транскрипционных факторов и других регуляторных белков. Некоторые последовательности ДНК не способны наматываться на нуклеосомные глобулы. Это последовательности, обладающие пониженной гибкостью, и последовательности, склонные с созданию неканонических структур, например, шпилек.
Из рисунка, представляющего скриншот UCSC, видна колокализация сайта ДНКазной гиперчувствительности (англ. DNaseI Hypersensitivity Clusters) с промоторами двух генов. Области открытого хроматина окружены , ацетилироваными по 27-му остатку лизина (H3K27Ac), что является меткой активных регуляторных областей хроматина, таких как промоторы и энхансеры. Кроме того, в районе сайта ДНКазной гиперчувствительности находится сайт связывания многих транскрипционных факторов, среди которых можно обнаружить консервативный фактор инициации транскрипции (является основной частью ). Также можно заметить частое связывание в этом районе РНК-полимеразы II, осуществляющей транскрипцию белок-кодирующих генов у человека. Для данного сайта ДНКазной гиперчувствительности характерна повышенная консервативность среди млекопитающих (англ. Mammal Cons), что означает сохранение этой последовательности в ходе эволюции, и, как следствие, её функциональное значение.
Ремоделирование хроматина
Образование областей, свободных от нуклеосом, происходит под действием специальных факторов, осуществляющих сборку, разборку и перемещение нуклеосом. Процесс изменения положения нуклеосом называется ремоделированием хроматина. В нём участвуют комплексы ремоделирования хроматина — консервативные белковые комплексы, работающие с затратой энергии АТФ. Ремоделирование хроматина осуществляется после внесения определенных эпигенетических меток — или . Если метки соответствуют активному хроматину (например, ацетилированый 9-й лизин гистона H3, ди- и триметилированный 4-й лизин гистона H3 и многие другие), то образуются участки открытого хроматина. Часто профиль модификаций гистонов имеет определенное распределение вокруг сайта гиперчувствительности к ДНКазе I.
Функциональные особенности эухроматина
Подобный вид хроматина рабочий и функционально активен. Не обладает свойством окрашивания и не определяется при гистологических исследованиях. В фазе митоза эухроматин практически весь конденсируется и становится составляющей частью хромосомы. Синтетические функции в этот период хромосомы не выполняют. Поэтому клеточные хромосомы могут находиться в двух функционально-структурных состояниях:
- Активное или рабочее состояние. В это время хромосомы почти полностью или полностью деконденсированы. Они участвуют в процессе транскрипции и редупликации. Все перечисленные процессы происходят непосредственно в ядре клетки.
- Неактивное состояние метаболического покоя (нерабочее). В этом состоянии хромосомы по максимуму конденсированы и служат транспортом для переноса генетического материала в дочерние клетки. В этом состоянии генетический материал еще и распределяется.
В завершающей фазе митоза происходит деспирализация и образуются слабоокрашенные структуры в виде нитей, содержащие транскрибируемые гены.
В структуре каждой хромосомы свой, уникальный, вариант расположения хроматина: эухроматина и гетерохроматина. Эта особенность клеток позволяет цитогенетикам идентифицировать отдельные хромосомы.
Типовые различия
Хроматин, в отличие от хромосом, способен существовать в двух типовых разновидностях:
Гетерохроматин -представляет собой хромосомы с очень низким или полностью отсутствующим уровнем функциональной активности. Гетерохроматин может быть и в виде частей хромосом. Он характеризуется относительной конденсированностью, которая представлена не в полном объёме. Под воздействием светового микроскопа гетерохроматин визуализируется как глыбки тёмного оттенка.
Зухроматин имеет характеристики, противоположные имеющимся характеристикам гетерохроматина. Он представляет собой хромосомы или их частичную фрагментацию, отличающуюся неполной степенью деконденсированности. С точки зрения функциональности эухроматин активен. Световой уровень не выявляет эухроматин, он продолжает даже под его воздействием оставаться практически неокрашенным.
Отличительной особенностью хроматина по сравнению с хромосомами можно считать и способность одного из его типов, гетерохроматина, в свою очередь делиться на два структурных элемента:
- Факультативный гетерохроматин, который отличает возможность трансформироваться в эухроматин.
- Конструктивный тип гетерохроматина, который не способен ни в какой из клеток осуществлять сходное преобразование.
Хромосомы и хроматин можно различать ещё и по внешнему облику. Хроматин представлен глыбками, гранулированными элементами и нитчатыми структурами, в целом можно сказать, что это уплотнённые участки хромосом.
Хромосомы, будучи единицей генетического материала, наделены плотной структурой.
Различна реакция хромосомы и хроматина на окрашивание. Хроматин поддаётся воздействию только некоторых красителей, к ним можно отнести кармин и гематоксилин. Хромосомы окрашиваются интенсивно, чутко реагируя на красящий элемент.





Строение гетерохроматина
Гетерохроматин имеет несложное строение (см. рисунок).

Гетерохроматин упаковывается в глобулы, которые называются нуклеосомами. Нуклеосомы образуют еще более плотные структуры и таким образом «мешают» считывать информацию с ДНК. Гетерохроматин образуется в процессе метилирования гистона НЗ по лизину 9, в последующем ассоциируется с белком 1 (НР1- Heterochromatin Protein 1). Также взаимодействует с другими белками, в том числе НЗК9-метилтрансфераз. Такое большое количество взаимодействий белков между собой является условием поддержания гетерохроматина и его распространения. Первичное строение ДНК не влияет на образование гетерохроматина.
Гетерохроматин — это не только отдельные части, но и целые хромосомы, которые на протяжении всего клеточного цикла остаются в конденсированном состоянии. Именно они в S-фазе и подвергаются репликации. Ученые считают, что гетерохроматиновые участки не несут в себе гены, которые кодируют белок, либо количество таких генов очень незначительно. Вместо таких генов нуклеотидные последовательности гетерохроматина в большинстве своих состоят из простых повторов.
Отличие хромосомы от хроматина
Различия на уровне структуры
Хромосомы, как элементы ядра клетки, имеют следующие отличительные особенности, расходящиеся с особенностями хроматина:
- Способность к самовоспроизведению.
- Наличие индивидуальной составляющей, связанной со структурными и функциональными свойствами.
- Возможность сохранять индивидуальные характеристики в ряду нескольких поколений.
Хроматин от хромосомы можно отличить по следующим параметрам:
- Является комплексом, переставленным сочетанием РНК, ДНК и белков.
- Представляет собой вещество хромосом.
- Располагается внутри клеток эукариот.
- Является составным фрагментом нуклеотида у прокариот.
- С помощью своего состава наделён способностью реализовывать генетическую информацию.
- Реализует репликацию и репарацию ДНК.
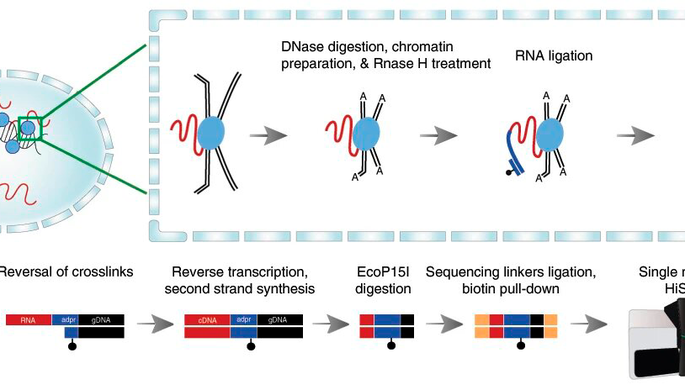
Картирование областей открытого хроматина
Для картирования областей открытого хроматина используют методы ДНКазной чувствительности (англ. DNase I hypersensitive) и изоляции регуляторных элементов с помощью формальдегида англ. formaldehyde-assisted isolation of regulatory elements (FAIRE). Метод ДНКазной чувствительности не позволяет определить, каким именно регуляторным участком является данная область открытого хроматина.
Ранее анализ результатов метода ДНКазной чувствительности проводился с помощью саузерн-блот гибридизации (англ. Southern blot). Это не позволяло проводить анализ большого количества сайтов, а также находить новые сайты гиперчувствительности. Анализ ДНКазной чувствительности можно проводить также с помощью ПЦР в реальном времени (количественной ПЦР). Это значительно проще, чем саузерн-блот гибридизация, но этот метод также имеет ограничение по количеству сайтов для анализа и не может быть использован для полногеномного исследования распределения сайтов чувствительности к ДНКазе I.
Развитие методов высокоэффективного секвенирования (англ. High-throughput sequencing) и ДНК-микрочипов (англ. DNA-microarray) позволяет картировать области открытого хроматина на всем протяжении генома. Кроме того, сочетание метода ДНКазной чувствительности с методом иммунопреципитации (англ. Chromatin immunoprecipitation (ChIP)) с последующим высокоэффективным секвенированием позволяет получать больше информации о связывании конкретных транскрипционных факторов с активными участками хроматина.
Другой способ картирования областей открытого хроматина — проведение иммунопреципитации хроматина (англ. ChIP) на антитела к гистонам. При этом области открытого хроматина должны быть мало представлены, так как с ними не связаны нуклеосомы. Метод ДНКазной чувствительности и иммунопреципитация гистонов дают сходные результаты.
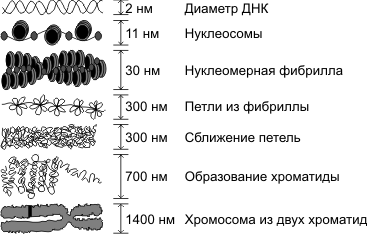
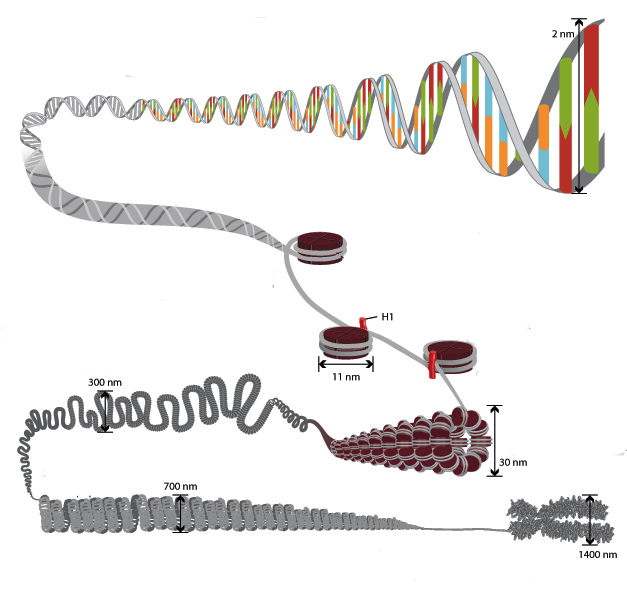
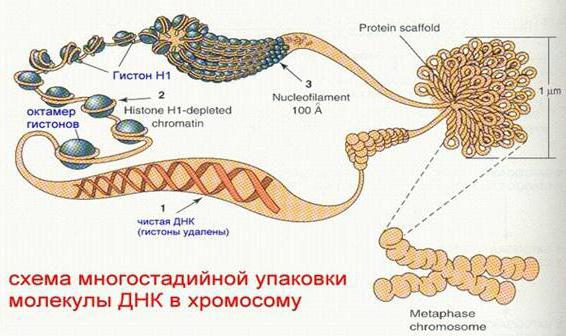
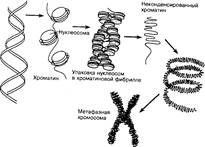
Уровни компактизации ДНК
Особенность структуры хроматина такая, что позволяет нитям ДНК с суммарной длиной более метра поместиться в ядро диаметром около 10 мкм. Такое возможно благодаря многоступенчатой системе упаковки генетических молекул. Общая схема компактизации включает пять уровней:
- нуклеосомная нить диаметром 10–11 нм;
- фибрилла 25–30 нм;
- петлевые домены (300 нм);
- волокно толщиной 700 нм;
- хромосомы (1200 нм).
Такая форма организации обеспечивает уменьшение длины исходной молекулы ДНК в 10 тысяч раз.

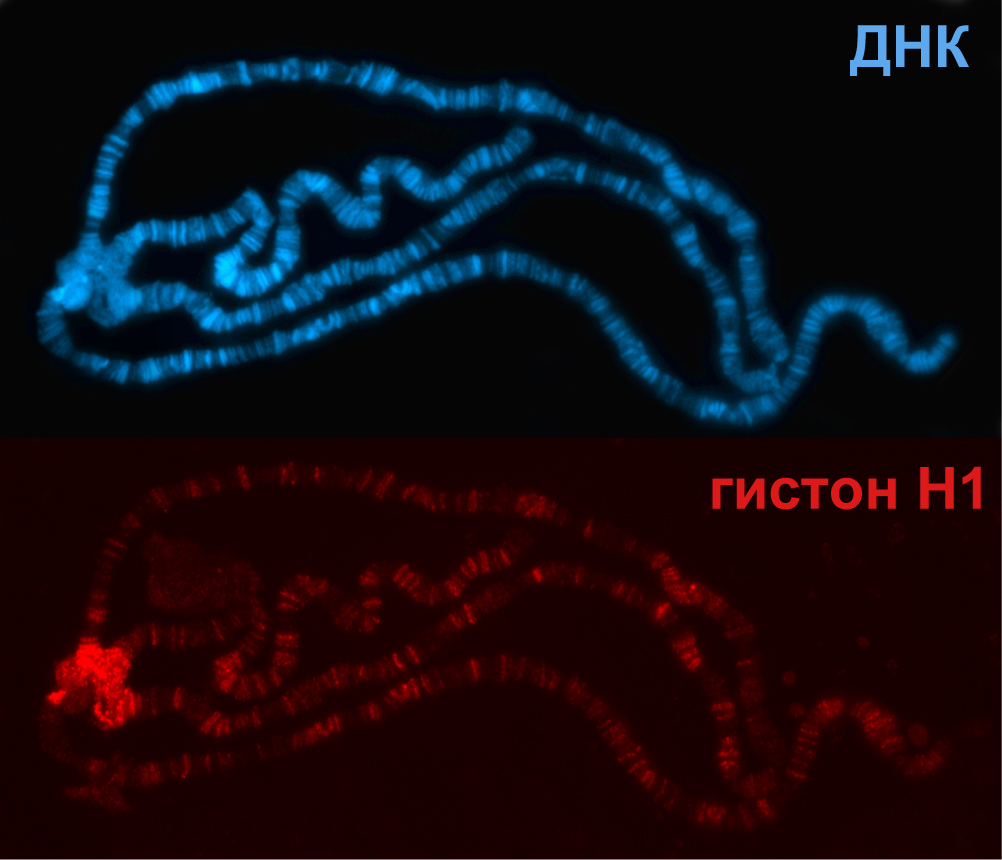
Нить диаметром 11 нм образована рядом нуклеосом, связанных линкерными участками ДНК. На электронной микрофотографии такая структура напоминает нанизанные на леску бусы. Нуклеосомная нить сворачивается в спираль по типу соленоида, образуя фибриллу толщиной 30 нм. В ее формировании участвует гистон H1.
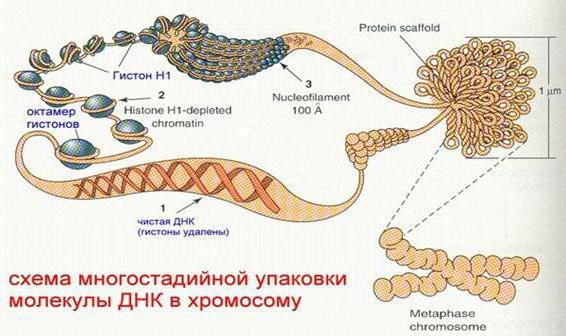

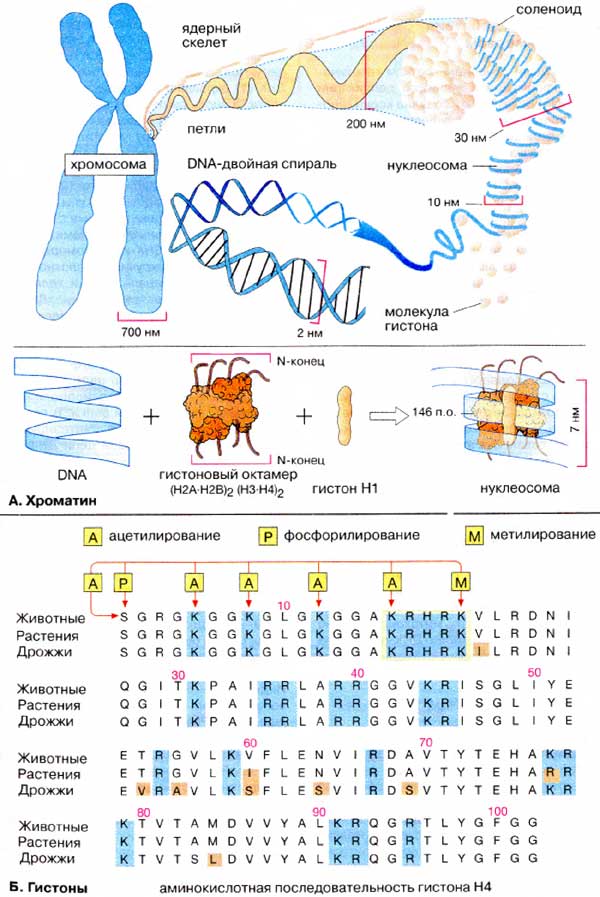


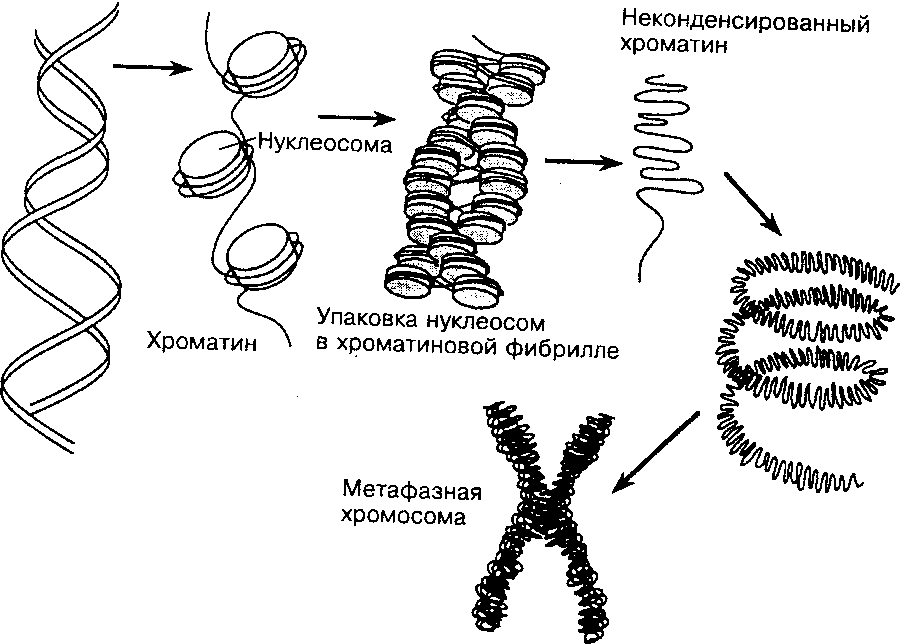


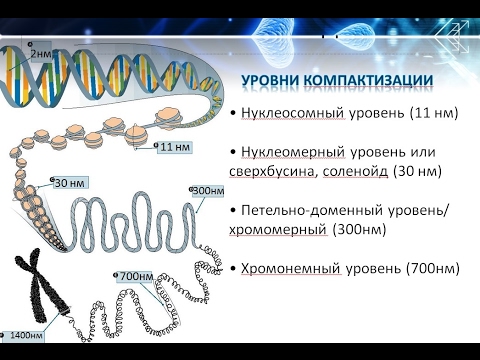
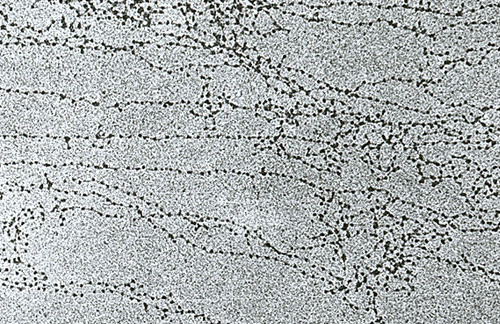
Соленоидная фибрилла складывается в петли (иначе – домены), которые закрепляются на поддерживающем внутриядерном матриксе. Каждый домен содержит от 30 до 100 тысяч пар нуклеотидов. Такой уровень компактизации характерен для интерфазного хроматина.
Структура толщиной 700 нм образуется при спирализации доменной фибриллы и называется хроматидой. В свою очередь, две хроматиды формируют пятый уровень организации ДНК – хромосому диаметром 1400 нм, которая становится видна на стадии митоза или мейоза.
Таким образом, хроматин и хромосома – это формы упаковки генетического материала, зависящие от жизненного цикла клетки.
От чего зависит конденсация вещества наследственности
Продолжая изучать вопрос «что такое хроматин», ученые установили, что его уплотнение зависит от белков-гистонов, входящих наряду с молекулами ДНК и РНК в состав нуклеосом. Они состоят из белков четырёх видов, называемых коровыми и линкерными. В момент транскрипции (считывание информации с генов с помощью РНК) вещество наследственности слабо конденсировано и носит название эухроматина.
В настоящее время особенности распределения молекул ДНК, связанных с гистоновыми белками, продолжают изучаться. Например, ученые выяснили, что хроматин различных локусов одной и той же хромосомы отличается уровнем конденсации. Например, в местах прикрепления к хромосоме нитей веретена деления, называемых центромерами, он более плотный, чем в теломерных участках – концевых локусах.
Восстановление хроматина и ДНК
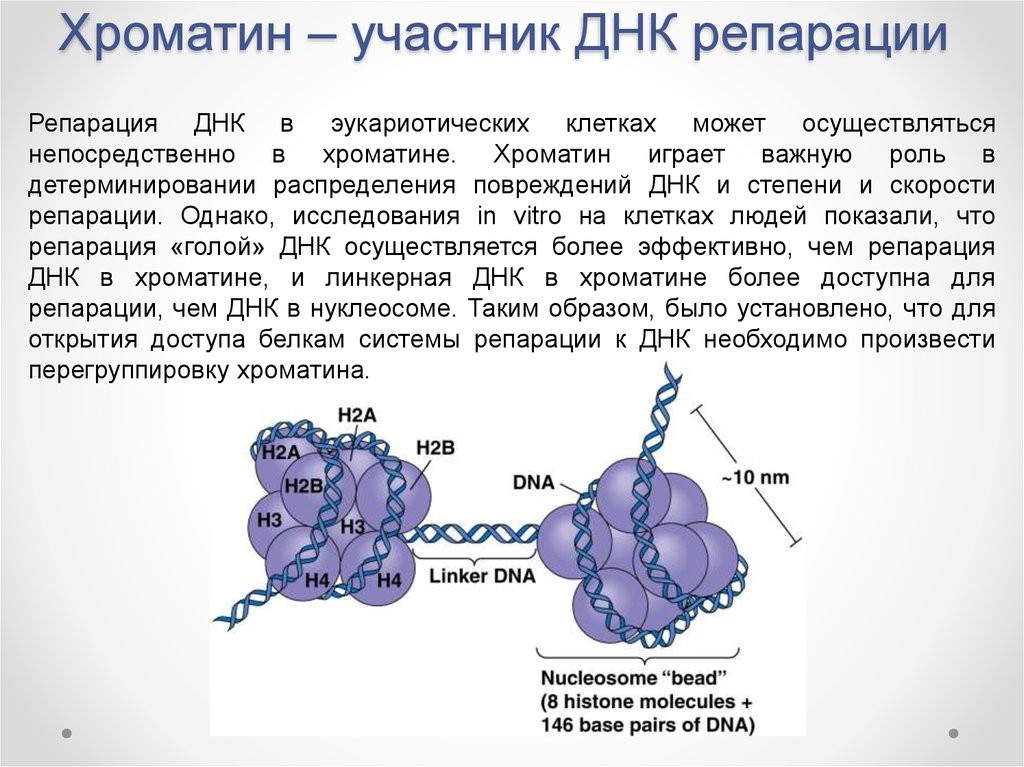
Упаковка эукариотической ДНК в хроматин представляет собой барьер для всех основанных на ДНК процессов, которые требуют привлечения ферментов к участкам их действия. Чтобы разрешить критический клеточный процесс репарации ДНК, хроматин должен быть реконструирован. У эукариот АТФ-зависимые комплексы ремоделирования хроматина и модифицирующие гистоны ферменты являются двумя преобладающими факторами, используемыми для осуществления этого процесса ремоделирования.
Релаксация хроматина происходит быстро в месте повреждения ДНК. Этот процесс инициируется белком PARP1, который начинает появляться при повреждении ДНК менее чем за секунду, с половинным максимальным накоплением в течение 1,6 секунды после повреждения. Затем ремоделирующий хроматин Alc1 быстро прикрепляется к продукту PARP1 и завершает прибытие к повреждению ДНК в течение 10 секунд после повреждения. Примерно половина максимальной релаксации хроматина, предположительно из-за действия Alc1, происходит через 10 секунд. Затем это позволяет задействовать фермент репарации ДНК MRE11 , чтобы инициировать репарацию ДНК в течение 13 секунд.
γH2AX, фосфорилированная форма H2AX , также участвует в ранних стадиях, ведущих к деконденсации хроматина после возникновения повреждения ДНК. Вариант гистона H2AX составляет около 10% гистонов H2A в хроматине человека. γH2AX (H2AX, фосфорилированный по серину 139) может быть обнаружен уже через 20 секунд после облучения клеток (с образованием двухцепочечного разрыва ДНК), а половина максимального накопления γH2AX происходит за одну минуту. Размер хроматина с фосфорилированным γH2AX составляет около двух миллионов пар оснований в месте двухцепочечного разрыва ДНК. Сам по себе γH2AX не вызывает деконденсацию хроматина, но в течение 30 секунд после облучения белок RNF8 может быть обнаружен в ассоциации с γH2AX. RNF8 опосредует обширную деконденсацию хроматина через его последующее взаимодействие с CHD4 , компонентом ремоделирования нуклеосом и деацетилазного комплекса NuRD .
После релаксации после повреждения ДНК с последующей репарацией ДНК хроматин восстанавливается до состояния уплотнения, близкого к уровню до повреждения, примерно через 20 мин.
Нобелевские премии
Следующие ученые были отмечены Нобелевскими премиями за их вклад в исследования хроматина :
| Год | ВОЗ | Награда |
|---|---|---|
| 1910 г. | Альбрехт Коссель (Гейдельбергский университет) | Нобелевская премия по физиологии и медицине за открытие пяти ядерных оснований: аденина , цитозина , гуанина , тимина и урацила . |
| 1933 г. | Томас Хант Морган (Калифорнийский технологический институт) | Нобелевская премия по физиологии и медицине за открытие роли гена и хромосомы в наследственности, основанное на его исследованиях белоглазой мутации у плодовой мухи Drosophila . |
| 1962 г. | Фрэнсис Крик , Джеймс Уотсон и Морис Уилкинс (Лаборатория молекулярной биологии MRC, Гарвардский и Лондонский университет соответственно) | Нобелевская премия по физиологии и медицине за открытие двойной спиральной структуры ДНК и ее значения для передачи информации в живом материале. |
| 1982 г. | Аарон Клаг (Лаборатория молекулярной биологии MRC) | Нобелевская премия по химии «за развитие кристаллографической электронной микроскопии и определение структуры биологически важных комплексов нуклеиновая кислота-белок» |
| 1993 г. | Ричард Дж. Робертс и Филип А. Шарп | Нобелевская премия по физиологии «за независимые открытия расщепленных генов », в которых участки ДНК, называемые экзонами, экспрессируют белки и прерываются участками ДНК, называемыми интронами , которые не экспрессируют белки. |
| 2006 г. | Роджер Корнберг (Стэнфордский университет) | Нобелевская премия по химии за открытие механизма, с помощью которого ДНК транскрибируется в информационную РНК. |
Локализация вещества наследственности в клетке
ДНК присутствует в таких цитоструктурах, как ядро, а также в органеллах, способных к делению — митохондриях и хлоропластах. Это связано с тем, что данные органоиды выполняют важнейшие функции в клетке: синтез АТФ, а также синтез глюкозы и образование кислорода в клетках растений. На синтетической стадии жизненного цикла материнские органеллы удваиваются. Таким образом, дочерние клетки в результате митоза (деления соматических клеток) или мейоза (образования яйцеклеток и сперматозоидов) получают необходимый арсенал клеточных структур, обеспечивающих клетки питательными веществами и энергией.

Рибонуклеиновая кислота состоит из одной цепи и имеет меньшую молекулярную массу, чем ДНК. Она содержится как в ядре, так и в гиалоплазме, а также входит в состав многих клеточных органоидов: рибосом, митохондрий, эндоплазматической сети, пластид. Хроматин в этих органеллах связан с белками-гистонами и входит в состав плазмид – кольцевых замкнутых молекул ДНК.


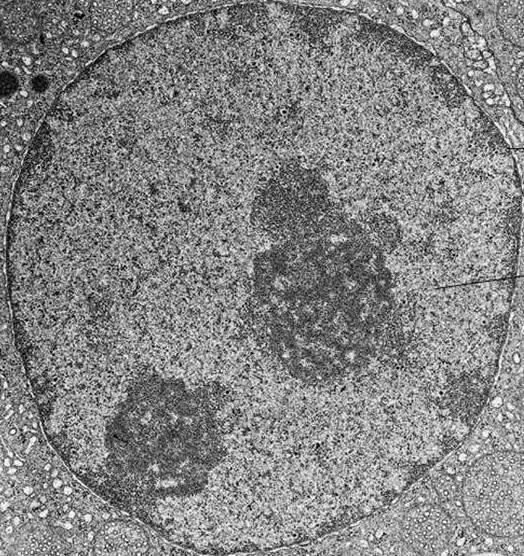



Эухроматин и гетерохроматин
Хроматин внутри клетки может быть уплотнен в различной степени в зависимости от стадии клетки в клеточном цикле. Хроматин в ядре содержится в виде эухроматина или гетерохроматина. Во время интерфазы, клетка не делится, а подвергается периоду роста. Большая часть хроматина находится в менее компактной форме, известной как эухроматин.
ДНК подвергается воздействию эухроматина, что позволяет проводить репликацию и транскрипцию ДНК. Во время транскрипции двойная спираль ДНК разматывается и открывается, чтобы можно было скопировать гены, кодирующие белки. Репликация и транскрипция ДНК необходимы для того, чтобы клетка синтезировала ДНК, белки и органеллы при подготовке к делению клеток (митоз или мейоз).
Небольшой процент хроматина существует как гетерохроматин во время интерфазы. Этот хроматин плотно упакован, что не позволяет проводить транскрипцию гена. Гетерохроматин окрашивается красителями в более темный цвет, чем эухроматин.
Хроматин и узлы
Было загадкой, как деконденсированные интерфазные хромосомы остаются по существу незаузленными. Естественное ожидание состоит в том, что в присутствии топоизомераз ДНК типа II, которые допускают прохождения двухцепочечных участков ДНК друг через друга, все хромосомы должны достичь состояния топологического равновесия. Топологическое равновесие в сильно переполненных интерфазных хромосомах, образующих хромосомные территории, должно приводить к образованию сильно узловатых волокон хроматина. Однако методы захвата конформации хромосом (3C) показали, что разрушение контактов с геномным расстоянием в интерфазных хромосомах практически такое же, как в смятом состоянии глобулы, которое образуется, когда длинные полимеры конденсируются без образования каких-либо узлов. Чтобы удалить узлы из сильно переполненного хроматина, потребуется активный процесс, который должен не только обеспечивать энергию для вывода системы из состояния топологического равновесия, но также направлять опосредованные топоизомеразой отрывки таким образом, чтобы узлы были эффективно развязаны вместо делая узлы еще более сложными. Было показано, что процесс экструзии петли хроматина идеально подходит для активного развязывания волокон хроматина в интерфазных хромосомах.
Ссылки
Дополнительные источники
- Cremer, T. 1985. Von der Zellenlehre zur Chromosomentheorie: Naturwissenschaftliche Erkenntnis und Theorienwechsel in der frühen Zell- und Vererbungsforschung, Veröffentlichungen aus der Forschungsstelle fürgerefische Patologie. Springer-Vlg., Берлин, Гейдельберг.
- Элгин, SCR (ред.). 1995. Структура хроматина и экспрессия генов, vol. 9. IRL Press, Оксфорд, Нью-Йорк, Токио.
- Поллард Т. и У. Эрншоу. 2002. Клеточная биология. Сондерс.
- Сомвебер, Х. 1987. Расположение хромосом в межфазных клеточных ядрах, стр. 223-234. В W. Hennig (ed.), Structure and Function of Eucaryotic Chromosomes, vol. 14. Шпрингер-Верлаг, Берлин, Гейдельберг.
- Ван Холде К.Э. 1989. Хроматин. Нью-Йорк: Springer-Verlag . ISBN 0-387-96694-3 .
- Ван Холде К., Дж. Златанова, Г. Аренц, Э. Мудрианакис. 1995. Элементы структуры хроматина: гистоны, нуклеосомы и волокна, с. 1-26. В SCR Elgin (ed.), Структура хроматина и экспрессия генов. IRL Press в издательстве Oxford University Press, Оксфорд.
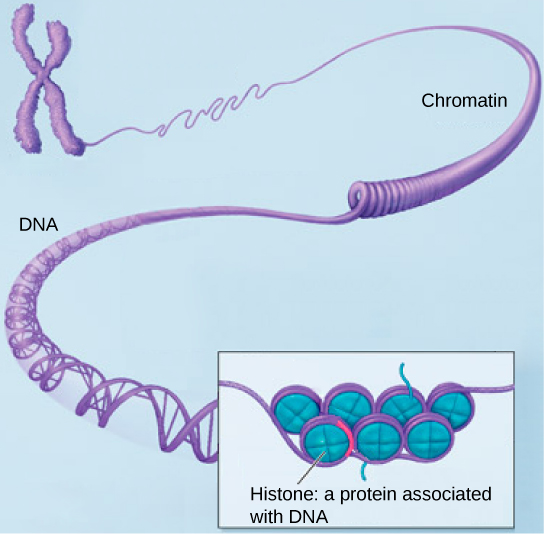
Хроматин, хромосома и хроматида
У людей часто возникают проблемы с различием терминов: хроматин, хромосома и хроматида. Хотя все три структуры состоят из ДНК и находятся внутри ядра, каждый из них определяется отдельно.
Хроматин состоит из ДНК и гистонов, которые упакованы в тонкие волокна. Эти волокна хроматина не конденсируются, но могут существовать либо в компактной форме (гетерохроматин), либо менее компактной форме (эухроматин). Процессы, включая репликацию ДНК, транскрипцию и рекомбинацию, встречаются в эухроматине. При делении клеток хроматин конденсируется с образованием хромосом.
Хромосомы представляют собой одноцепочечные структуры конденсированного хроматина. Во время процессов деления клеток через митоз и мейоз, хромосомы реплицируются, чтобы гарантировать, что каждая новая дочерняя клетка получает правильное количество хромосом. Дублицированная хромосома является двухцепочечной и имеет привычную форму X. Две нити идентичны и связаны в центральной области, называемой центромером.
Хроматидой является одна из двух нитей реплицированных хромосом. Хроматиды, соединенные центромером, называются сестринскими хроматидами. В конце клеточного деления сестринские хроматиды отделяются от дочерних хромосом в новообразованных дочерних клетках.
Хроматин и всплески транскрипции
Хроматин и его взаимодействие с ферментами были исследованы, и был сделан вывод, что он является важным фактором экспрессии генов. Винсент Г. Олфри, профессор Университета Рокфеллера, заявил, что синтез РНК связан с ацетилированием гистонов. Аминокислота лизин, прикрепленная к концам гистонов, заряжена положительно. Ацетилирование этих хвостов сделает концы хроматина нейтральными, что обеспечит доступ к ДНК.
Когда хроматин деконденсируется, ДНК открыта для проникновения молекулярных механизмов. Колебания между открытым и закрытым хроматином могут способствовать прерыванию транскрипции или взрыву транскрипции . Вероятно, вовлечены и другие факторы, такие как ассоциация и диссоциация комплексов факторов транскрипции с хроматином. Этот феномен, в отличие от простых вероятностных моделей транскрипции, может объяснять высокую вариабельность экспрессии генов между клетками в изогенных популяциях.
Альтернативные организации хроматина
Во время многоклеточного сперматогенеза , в сперматидном хроматине «сек ремоделируется в более разнесен упакованным, уширенном, почти кристаллоподобную структуру. Этот процесс связан с прекращением транскрипции и включает обмен ядерными белками. Гистоны в основном вытесняются и заменяются протаминами (маленькими белками, богатыми аргинином ). Предполагается, что у дрожжей области, лишенные гистонов, становятся очень хрупкими после транскрипции; HMO1, белок HMG-box , помогает в стабилизации хроматина, свободного от нуклеосом.
Гены-регуляторы и состав хроматина
В концепции регуляции генной активности, созданной французскими генетиками Жакобом и Моно, дается представление о существовании участков дезоксирибонуклеиновой кислоты, в которых нет информации о структурах белков. Они выполняют чисто бюрократические – управленческие функции. Называясь генами-регуляторами, эти части хромосом, как правило, в своей структуре лишены белков-гистонов. Хроматин, определение которого было проведено методом секвенирования, получил название открытого.
В ходе дальнейших исследований было установлено, что в этих локусах расположены последовательности нуклеотидов, препятствующие присоединению к молекулам ДНК белковых частиц. Такие участки содержат регуляторные гены: промоторы, эхансеры, активаторы. Компактизация хроматина в них высока, а длина этих участков в среднем составляет около 300 нм. Существует биохимический метод определения открытого хроматина в изолированных ядрах, при котором используют фермент ДНК-азу. Он очень быстро расщепляет локусы хромосом, лишенные белков-гистонов. Хроматин в этих участках был назван сверхчувствительным.






Понятие хромосомы и хроматина
Хромосомой принято называть элемент, входящий в состав ядра клетки. Хромосома участвует в формировании структуры ядра. Это хранилище ДНК, а значит и информации наследственного типа об организме в целом. Линейный порядок расположения генов характеризует хромосомы. Хромосому формируют хроматиды. Хроматиды представлены при этом парой продольных субЪединиц. Каждая из входящих в обозначенную пару хроматид по своему строению и структуре абсолютно аналогична другой хроматиде. Основу хроматиды составляет молекула ДНК, представленная в единичном экземпляре. Теломеры выступают конечными участками хроматид.

Хромосомы
Хроматин является веществом, входящим в состав хромосомы. Его можно выделить из ядер клеток растений или животных. Для хроматина характерна способность интенсивно окрашиваться ядерными красителями. Когда клетка начинает делиться, хроматин претерпевает процесс формирования в различимые структуры определённого типа, находящиеся в составе хромосом.
Хроматин: альтернативные определения
Термин, введенный Вальтером Флеммингом , имеет несколько значений:
- Простое и краткое определение: хроматин — это макромолекулярный комплекс макромолекулы ДНК и макромолекул белка (и РНК). Белки упаковывают и упорядочивают ДНК и контролируют ее функции в ядре клетки.
- Оперативное определение биохимиков: Хроматин — это комплекс ДНК / белок / РНК, извлеченный из лизированных межфазных ядер эукариот. Какое из множества веществ, присутствующих в ядре, будет составлять часть извлеченного материала, частично зависит от метода, который использует каждый исследователь. Кроме того, состав и свойства хроматина варьируются от одного типа клеток к другому, во время развития определенного типа клеток и на разных стадиях клеточного цикла.
- Определение ДНК + гистон = хроматин : двойная спираль ДНК в ядре клетки упакована специальными белками, называемыми гистонами. Образованный комплекс белок / ДНК называется хроматином. Основная структурная единица хроматина — нуклеосома.
Первое определение позволяет определять «хроматины» в других областях жизни, таких как бактерии и археи, с использованием любых ДНК-связывающих белков, которые конденсируют молекулу . Эти белки обычно относят к (NAP); примеры включают AsnC / LrpC с HU. Кроме того, некоторые археи действительно производят нуклеосомы из белков, гомологичных эукариотическим гистонам.