Мутации
Они происходят в среднем с частотой 10-5 на ген на гамету на поколение. Все аллели, которые обнаруживаются в группах, когда-то возникли вследствие мутации. Чем меньше популяция, тем ниже вероятность того, что каждое поколение будет иметь хотя бы одну особь — носителя новой мутации. При численности в сто тысяч у каждой группы потомков с вероятностью, приближенной к единице, найдется мутантный аллель. Однако его частота в популяции, а также возможность его закрепления будет достаточно низкой. Вероятность, что эта же мутация появится в том же поколении хотя бы у одной особи при численности 10, ничтожно мала. Однако если она все же произойдет в данной популяции, то частота мутантного аллеля (1 на 20 аллелей), а также шансы на его фиксацию будут относительно высокими. В больших популяциях возникновение нового элемента происходит относительно быстро. При этом его закрепление проходит медленно. Малочисленные популяции, напротив, мутацию ожидают долго. Но после ее возникновения закрепление проходит быстро. Из этого можно сделать следующий вывод: шанс на фиксацию нейтральных аллелей находится в зависимости только от частоты мутационного возникновения. При этом численность популяции на этот процесс не влияет.

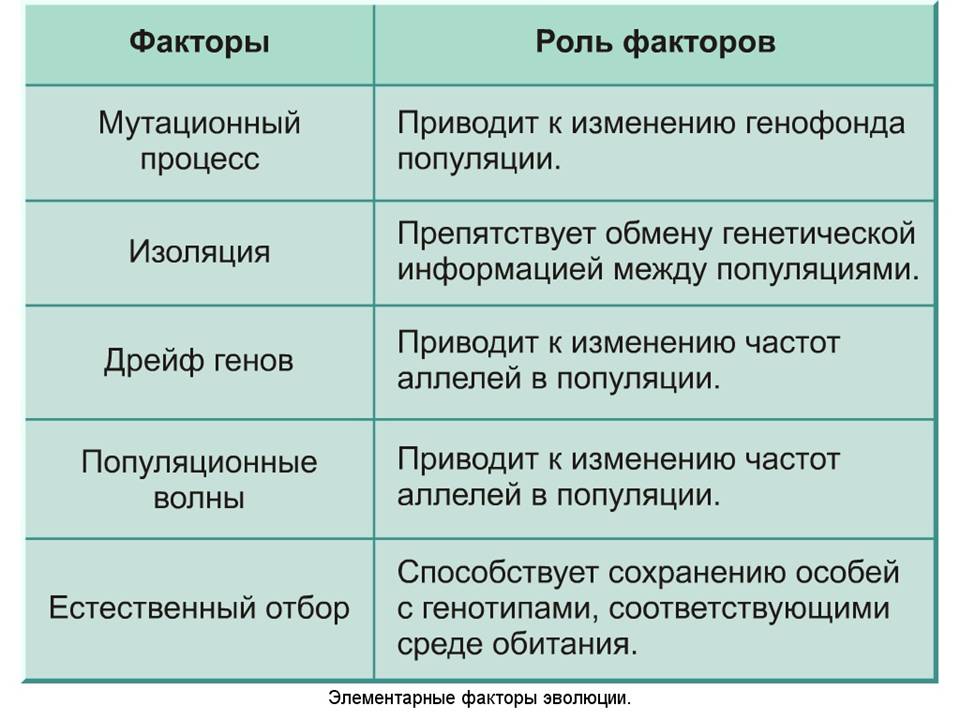
Соотношение влияния
В процессе развития генетический дрейф взаимодействует с прочими факторами. Прежде всего, взаимосвязь устанавливается с естественным отбором. Соотношение вкладов данных факторов находится в зависимости от ряда обстоятельств. В первую очередь его определяет интенсивность отбора. Вторым обстоятельством является численность группы. Так, если интенсивность и численность высоки, случайные процессы обладают ничтожно малым влиянием на динамику генетических частот. При этом в небольших группах при несущественных различиях в приспособленности влияние изменений несравнимо больше. В таких случаях возможна фиксация менее адаптивного аллеля, при том что более адаптивный будет потерян.
Биология
Учебник для 10-11 классов
§ 48. Дрейф генов — фактор эволюции
Случайные колебания частот генов в популяциях ограниченного размера. Случайные отклонения результатов расщеплений при моно-гибридном, дигибридном и других типах скрещиваний от ожидаемых величин — явление обычное. Даже в опыте Г. Менделя во втором поколении соотношение желтых семян к зеленым составило 6022:2001, т. е. не было точно равно 3:1. Если бы в подобном опыте было изучено не 8000 семян, а только 80, то вероятность получить соотношение 3:1 была бы существенно ниже. В малых популяциях действие случайных процессов приводит к заметным последствиям, в частности к изменениям частот аллелей. Случайное ненаправленное изменение частот аллелей в популяции получило название дрейфа генов.
Явление генетического дрейфа впервые обнаружили российские генетики Н. П. Дубинин и Д. Д. Ромашов, а также зарубежные ученые С. Райт и Р. Фишер. С. Райт экспериментально доказал, что в маленьких популяциях частота мутантного аллеля меняется быстро и случайным образом. Его опыт был прост: в пробирки с кормом он посадил по две самки и два самца мух дрозофил, гетерозиготных по гену А (их генотип можно записать Аа). В этих искусственно созданных популяциях концентрация нормального (А) и мутантного (а) аллелей составила 50%. Через несколько поколений оказалось, что в некоторых популяциях все особи стали гомозиготными по мутантному аллелю (а), в других популяциях он был вовсе утрачен, и, наконец, часть популяций содержала как нормальный, так и мутантный аллель
Важно подчеркнуть, что, несмотря на снижение жизнеспособности мутантных особей и, следовательно, вопреки естественному отбору, в некоторых популяциях мутантный аллель полностью вытеснил нормальный. Это и есть результат случайного процесса — дрейфа генов
Популяционные волны. В природных условиях периодические колебания численности различных организмов очень распространены. На рисунке 61 в качестве примера показаны изменения численности популяции хищника и жертвы. Видно, что в разные годы происходит резкое возрастание и падение численности животных, причем изменения численности жертвы как бы опережают численность хищника. С. С
Четвериков одним из первых обратил внимание на периодические колебания численности популяции. Колебания численности особей, составляющих популяцию, получили название популяционных волн.
Рис. 61. Популяционные волны (динамика численности зайцев и хищников)










Популяционные волны — одна из частых причин дрейфа генов. Особенно сильно колебания численности выражены у насекомых, размер весенней популяции у которых обычно сокращается в тысячи раз по сравнению с осенними популяциями. Случайное выживание редких мутантных особей в период зимовки может увеличить концентрацию данной мутации в тысячи раз.
К каким последствиям для популяции приводит дрейф генов? Они могут быть различными. Во-первых, может возрастать генетическая однородность популяции, т. е. возрастает ее гомозиготность. Кроме того, популяции, имеющие вначале сходный генетический состав и обитающие в сходных условиях, могут в результате дрейфа различных генов утратить первоначальное сходство. Во-вторых, вследствие дрейфа генов, вопреки естественному отбору, в популяции может удерживаться аллель, снижающий жизнеспособность особей. И наконец, в-третьих, благодаря популяционным волнам может происходить быстрое и резкое возрастание концентраций редких аллелей.
Таким образом, можно сказать, что дрейф генов в популяции возникает в результате различных случайных процессов и вносит вклад в эволюционные преобразования генотипической структуры популяций.
- Объясните, что такое дрейф генов. Приведите пример ситуации, в которой он играет важную роль, и объясните, почему его роль особенно велика в небольших популяциях.
- Какую роль играют в эволюции дрейф генов и популяционные волны?
примеров
Гипотетический пример: улитки и коровы
Представьте себе луг, где сосуществуют улитки и коровы. В популяции улиток можно выделить две окраски: черная и желтая. Решающим фактором смертности улиток являются следы коров.
Однако обратите внимание, что если на улитку наступают, это не зависит от цвета ее раковины, так как это случайное событие. В этом гипотетическом примере популяция улиток начинается с равной пропорции цветов (50 черных улиток и 50 желтых улиток)
В случае коров исключают 6 черных и только 2 желтых, соотношение обменных цветов.
Точно так же в следующем случае желтые могут погибнуть в большей пропорции, так как нет никакой связи между цветом и вероятностью раздавливания (однако, нет типа «компенсирующего» эффекта)..
Как изменится доля улиток с течением времени?
Во время этого случайного процесса пропорции черных и желтых оболочек будут колебаться с течением времени. В конце концов, одна из оболочек достигнет любого из двух пределов: 0 u 1.
Когда достигнутая частота равна 1 — предположим, что для аллели желтой раковины — все улитки будут такого цвета. И, как мы можем догадаться, аллель для черной раковины будет потерян.
Единственный способ получить этот аллель снова — это популяция через миграцию или мутацию..









Гены дрейфуют в действии: Гепарды
Явление дрейфа генов можно наблюдать в природных популяциях, и наиболее ярким примером являются гепарды. Эти быстрые и стильные кошачьи принадлежат к виду Acinonyx jubatus.
Около 10 000 лет назад гепарды и другие популяции крупных млекопитающих пережили крайнее исчезновение. Это событие вызвало «узкое место» в городе гепардов, где выжило лишь несколько человек.
Оставшиеся в живых после катастрофического явления плейстоцена породили всех гепардов современности. Эффекты дрейфа в сочетании с инбридингом почти полностью гомогенизируют население.
На самом деле, иммунная система этих животных практически одинакова у всех людей. Если по какой-либо причине кому-либо из членов понадобилось пожертвование какого-либо органа, любой из их партнеров мог бы сделать это, не приводя к шансу отказа.
Пожертвования — это процедуры, которые выполняются тщательно, и необходимо подавить иммунную систему получателя, чтобы она не атаковала «внешнего агента», даже если он исходит от очень близкого родственника — будь то братья или сыновья.
Пример в человеческих популяциях: амиши
Узкие места и основополагающий эффект также встречаются в современной человеческой популяции и имеют очень важные последствия в области медицины.
Амиши являются религиозной группой. Они характеризуются простым образом жизни, свободным от технологий и других современных удобств — в дополнение к чрезвычайно высокой частоте заболеваний и генетических патологий..
Около 200 колонизаторов прибыли в Пенсильванию (США) из Европы и начали размножаться среди тех же членов..
Предполагается, что среди колонизаторов были носители аутосомно-рецессивных генетических заболеваний, в том числе синдром Эллиса-ван Кревельда. Этот синдром характеризуется особенностями карликовости и полидактилии (большое количество пальцев, больше пяти цифр).
Заболевание было обнаружено в исходной популяции с частотой 0,001 и значительно возросло до 0,07..
Исследования Поллинга и Цукуркендла
Эти два американских ученых выявили, что число различий в аминокислотной последовательности в цитохроме и гемоглобине у тех или иных видов млекопитающих тем выше, чем раньше произошло расхождение их эволюционных путей. Впоследствии данная закономерность была подтверждена большим объемом экспериментальных данных. Материал включал в себя десятки различных генов и несколько сотен видов животных, микроорганизмов и растений. Выяснилось, что ход молекулярных часов осуществляется с постоянной скоростью. Это открытие, собственно говоря, подтверждается рассматриваемой теорией. Калибровка часов производится отдельно для каждого гена. Это обусловлено тем, что частота появления нейтральных мутаций у них различна. Для этого проводится оценка количества замен, скопившихся в определенном гене у таксонов. Их время дивергенции надежно установлено с помощью палеонтологических данных. После того как будут откалиброваны молекулярные часы, их можно использовать далее. В частности, с их помощью легко измерить время, в течение которого произошла дивергенция (расхождение) между различными таксонами. Это возможно даже в том случае, если общий их предок еще не выявлен в палеонтологической летописи.









Пояснение
Один из механизмов дрейфа генов заключается в следующем. В процессе размножения в популяции образуется большое число половых клеток — гамет. Большая часть этих гамет не формирует зигот. Тогда новое поколение в популяции формируется из выборки гамет, которым удалось образовать зиготы. При этом возможно смещение частот аллелей относительно предыдущего поколения.
Дрейф генов на примере
Механизм дрейфа генов может быть продемонстрирован на небольшом примере. Представим очень большую колонию бактерий, находящуюся изолированно в капле раствора. Бактерии генетически идентичны за исключением одного гена с двумя аллелями A и B. Аллель A присутствует у одной половины бактерий, аллель B — у другой. Поэтому частота аллелей A и B равна 1/2. A и B — нейтральные аллели, они не влияют на выживаемость или размножение бактерий. Таким образом, все бактерии в колонии имеют одинаковые шансы на выживание и размножение.
Затем размер капли уменьшаем таким образом, чтобы питания хватало лишь для 4 бактерий. Все остальные умирают без размножения. Среди четырёх выживших возможно 16 комбинаций для аллелей A и B:
(A-A-A-A), (B-A-A-A), (A-B-A-A), (B-B-A-A), (A-A-B-A), (B-A-B-A), (A-B-B-A), (B-B-B-A), (A-A-A-B), (B-A-A-B), (A-B-A-B), (B-B-A-B), (A-A-B-B), (B-A-B-B), (A-B-B-B), (B-B-B-B).
Вероятность каждой из комбинаций
- 12⋅12⋅12⋅12=116{\displaystyle {\frac {1}{2}}\cdot {\frac {1}{2}}\cdot {\frac {1}{2}}\cdot {\frac {1}{2}}={\frac {1}{16}}}
где 1/2 (вероятность аллеля A или B для каждой выжившей бактерии) перемножается 4 раза (общий размер результирующей популяции выживших бактерий)
Если сгруппировать варианты по числу аллелей, то получится следующая таблица:
| A | B | Количество вариантов | Вероятность |
| 4 | 1 | 1/16 | |
| 3 | 1 | 4 | 4/16 |
| 2 | 2 | 6 | 6/16 |
| 1 | 3 | 4 | 4/16 |
| 4 | 1 | 1/16 |
Как видно из таблицы, в шести вариантах из 16 в колонии будет одинаковое количество аллелей A и B. Вероятность такого события 6/16. Вероятность всех прочих вариантов, где количество аллелей A и B неодинаково несколько выше и составляет 10/16.
Дрейф генов происходит при изменении частот аллелей в популяции из-за случайных событий. В данном примере популяция бактерий сократилась до 4 выживших (эффект бутылочного горлышка). Сначала колония имела одинаковые частоты аллелей A и B, но шансы, что частоты изменятся (колония подвергнется дрейфу генов) выше, чем шансы на сохранение оригинальной частоты аллелей. Также существует высокая вероятность (2/16), что в результате дрейфа генов один аллель будет утрачен полностью.
Общие сведения
Дрейф генов, примеры которого будут приведены в статье далее, представляет собой определенные изменения, которые фиксируются от поколения к поколению. Считается, что у этого явления существуют свои собственные механизмы. Некоторые исследователи обеспокоены тем, что в генофонде многих, если не всех, наций в настоящий момент довольно быстро увеличивается объем появляющихся аномальных генов. Они определяют наследственную патологию, формируют предпосылки для развития множества других заболеваний. Считается также, что патоморфоз (изменение признаков) разных болезней, в том числе и болезней психического характера, обусловливает именно дрейф генов. Явление, о котором идет речь, происходит стремительными темпами. В итоге ряд психических расстройств принимает неизвестные формы, становятся неузнаваемыми при сопоставлении с их описанием в классических изданиях. Вместе с этим существенные изменения отмечаются и непосредственно в самой структуре психиатрической заболеваемости. Так, дрейф генов стирает некоторые формы шизофрении, встречавшиеся ранее. Вместо них появляются такие патологии, которые с трудом можно определить по современным классификаторам.
Дрейф и фиксация
Закон Харди — Вайнберга гласит, что в достаточно больших популяциях частоты аллелей остаются постоянными от поколения к поколению, если только равновесие не нарушено миграцией, генетическими мутациями или отбором.
Однако в конечных популяциях новые аллели не получены в результате случайной выборки аллелей, переданной следующему поколению, но выборка может привести к исчезновению существующего аллеля. Поскольку случайная выборка может удалить, но не заменить аллель, и поскольку случайное снижение или увеличение частоты аллелей влияет на ожидаемое распределение аллелей для следующего поколения, генетический дрейф ведет популяцию к генетической однородности с течением времени. Когда аллель достигает частоты 1 (100 %), она считается «фиксированной» в популяции, а когда аллель достигает частоты 0 (0 %), она исчезает. Меньшие популяции достигают фиксации быстрее, тогда как в пределе бесконечной популяции фиксация не достигается. Как только аллель становится фиксированной, генетический дрейф останавливается, и частота аллеля не может измениться, если новый аллель не введен в популяцию посредством мутации или потока генов. Таким образом, даже несмотря на то, что генетический дрейф является случайным, бесцельным процессом, он устраняет генетические изменения во времени.










Скорость изменения частоты аллелей из-за дрейфа
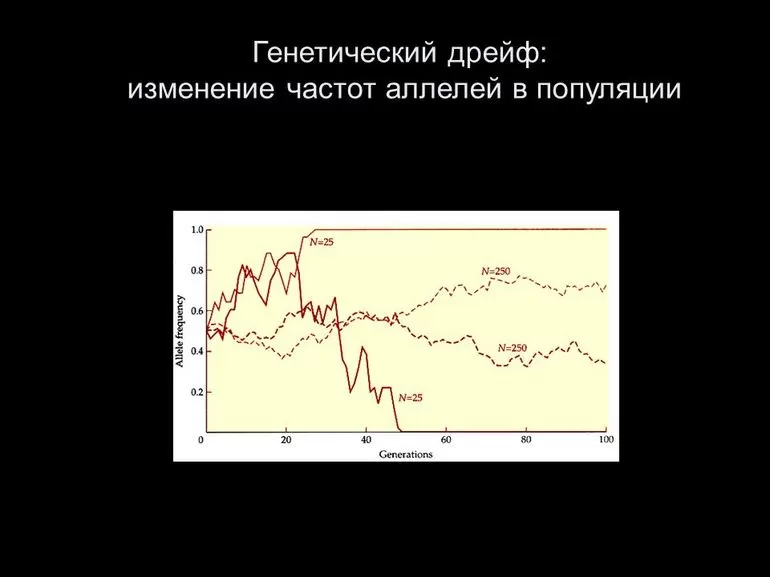
Десять симуляций случайного генетического дрейфа одного данного аллеля с исходным частотным распределением 0,5, измеренным в течение 50 поколений, повторенных в трех репродуктивно синхронных популяциях разных размеров. В этих моделях аллели дрейфуют к потере или фиксации (частота 0,0 или 1,0) только в самой маленькой популяции.
Предполагая, что генетический дрейф является единственной эволюционной силой, действующей на аллель, после t поколений во многих реплицируемых популяциях, начиная с частот аллелей p и q, дисперсия частоты аллелей между этими популяциями
- Vt≈pq(1−exp(−t2Ne)){\displaystyle V_{t}\approx pq\left(1-\exp \left(-{\frac {t}{2N_{e}}}\right)\right)}
Теория Райта
Случайный дрейф генов изучался с помощью математических моделей. Используя этот принцип, Райт вывел теорию. Он считал, что решающее значение дрейфа генов при постоянных условиях отмечается в небольших группах. Они становятся гомозиготными, и изменчивость уменьшается. Райт также полагал, что вследствие изменений в группах способны сформироваться негативные наследственные признаки. В результате этого вся популяция может погибнуть, не внеся вклад в развитие вида. Вместе с этим большую роль во многих группах играет отбор. В этой связи генетическая изменчивость внутри популяции вновь будет несущественной. Постепенно группа хорошо приспособится к окружающим условиям. Однако последующие эволюционные изменения будут зависеть от возникновения благоприятных мутаций. Эти процессы проходят достаточно медленно. В этой связи и эволюция больших популяций не отличается высокой скоростью. В группах промежуточной величины отмечается повышенная изменчивость. При этом образование новых выгодных генов происходит случайно, что, в свою очередь, ускоряет эволюцию.
Против естественного отбора
В природных популяциях генетический дрейф и естественный отбор не действуют изолированно, оба явления всегда играют роль, вместе с мутацией и миграцией. Нейтральная эволюция является продуктом как мутации, так и дрейфа, а не только дрейфа. Точно так же, даже когда отбор преодолевает генетический дрейф, он может действовать только на не нейтральные вариации.
В то время как естественный отбор имеет направление в сторону наследственных адаптаций к текущей среде, генетический дрейф не имеет направления и зависит только от математической случайности. В результате дрейф воздействует на (числа особей с одинаковым набором ДНК) в популяции, независимо от их фенотипических признаков. В отличие от этого, отбор способствует распространению аллелей, фенотипические эффекты которых увеличивают выживаемость и / или плодовитость их носителей, снижает частоту аллелей, вызывающих неблагоприятные признаки, и игнорирует нейтральные.
Закон больших чисел предсказывает, что, когда абсолютное число копий аллеля мало (например, в ), величина дрейфа на частотах аллеля на поколение больше. Величина дрейфа достаточно велика, чтобы подавить отбор при любой частоте аллеля, когда меньше 1, деленного на эффективный размер популяции. Поэтому считается, что не адаптивная эволюция, возникающая в результате мутации и генетического дрейфа, является последующим механизмом эволюционных изменений, главным образом в небольших изолированных популяциях. Математика генетического дрейфа зависит от эффективного размера популяции, но неясно, как это связано с фактическим числом особей в популяции. Генетическая связь с другими генами, которые находятся в процессе отбора, может уменьшить эффективный размер популяции, испытываемый нейтральным аллелем. При более высокой частоте рекомбинации уменьшается связь, и вместе с этим это локальное влияние на эффективный размер популяции. Этот эффект виден в молекулярных данных как корреляция между локальной скоростью рекомбинации и генетическим разнообразием и отрицательной корреляцией между плотностью и разнообразием генов в не кодирующих областях ДНК. Стохастичность, связанная со связью с другими генами, находящимися в процессе отбора, не является той же ошибкой, что и выборка, и иногда её называют генетической тягой, чтобы отличить её от генетического автостопа.
Когда частота аллелей очень мала, дрейф также может усиливать отбор даже в больших популяциях. Например, в то время как неблагоприятные мутации обычно быстро устраняются в больших популяциях, новые полезные мутации почти так же уязвимы для потери из-за генетического дрейфа, как и нейтральные мутации. Только когда частота аллеля для преимущественной мутации достигнет определённого порога, генетический дрейф не будет иметь никакого эффекта.
Литература
- Михеев В. С. . Большая российская энциклопедия (2007). Дата обращения 2 декабря 2019.
- Воронцов Н.Н., Сухорукова Л.Н. Эволюция органического мира. — М.: Наука, 1996. — С. 93—96. — ISBN 5-02-006043-7.
- Грин Н., Стаут У., Тейлор Д. Биология. В 3 томах. Том 2. — М.: Мир, 1996. — С. 287—288. — ISBN 5-03-001602-3.
- Hartl, Daniel L.; Clark, Andrew G. (англ.)русск.. Principles of Population Genetics (неопр.). — 4th. — Sunderland, MA: Sinauer Associates (англ.)русск., 2007. — ISBN 978-0-87893-308-2.
- Futuyma, Douglas (англ.)русск.. Evolutionary Biology (неопр.). — 3rd. — Sunderland, MA: Sinauer Associates (англ.)русск., 1998. — ISBN 0-87893-189-9.
- Ewens, Warren J. (англ.)русск.. Mathematical Population Genetics I. Theoretical Introduction (англ.). — 2nd. — New York: Springer Science+Business Media, 2004. — Vol. 27. — (Interdisciplinary Applied Mathematics). — ISBN 0-387-20191-2.
- Barton, Nicholas H. (англ.)русск.; Briggs, Derek E.G. (англ.)русск.; Eisen, Jonathan A. (англ.)русск.; Goldstein, David B. (англ.)русск.; Patel, Nipam H. Evolution (неопр.). — Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press (англ.)русск., 2007. — ISBN 978-0-87969-684-9.
- Non-Neutral Evolution: Theories and Molecular Data (англ.) / Golding, Brian. — New York: Chapman & Hall, 1994. — ISBN 0-412-05391-8. «Материалы семинара, спонсируемого Канадским институтом перспективных исследований».
Последствия изменений
Одним из основных результатов генетического дрейфа выступает обеднение разнообразия внутри группы. Это происходит за счет утраты одних аллелей и фиксации других. Процесс мутации, в свою очередь, напротив, способствует обогащению внутри популяций генетического разнообразия. За счет мутирования утерянный аллель может возникать снова и снова. В связи с тем что генетический дрейф представляет собой направленный процесс, одновременно со снижением внутрипопуляционного разнообразия увеличивается различие между локальными группами. Противодействует этому явлению миграция. Так, если в одной популяции фиксируется аллель «А», а в другой – «а», то внутри этих групп снова появляется разнообразие.
ссылки
- Одесирк Т., Одесирк Г. и Байерс Б.Э. (2004). Биология: наука и природа. Пирсон Образование.
- Кертис Х. & Шнек А. (2006). Приглашение к биологии. Ed. Panamericana Medical.
- Фриман, С. & Херрон, Дж. С. (2002). Эволюционный анализ. Прентис Холл.
- Футуйма, Д.Дж. (2005). эволюция . Sinauer.
- Hickman, C.P., Roberts, L.S., Larson, A., Ober, W.C., & Garrison, C. (2001). Интегрированные принципы зоологии (Том 15). Нью-Йорк: Макгроу-Хилл.
- Майр, Э. (1997). Эволюция и разнообразие жизни: избранные очерки. Издательство Гарвардского университета.
- Райс С. (2007).Энциклопедия Эволюции. Факты в файле.
- Рассел П., Герц П. и Макмиллан Б. (2013). Биология: Динамическая Наука. Нельсон Образование.
- Солер М. (2002). Эволюция: основа биологии. Южный Проект.
Выводы Райта
Когда из популяции теряется один аллель, появиться он может вследствие определенной мутации. Но если вид разделяется на несколько групп, в одной из которых отсутствует один элемент, в другой – другой, то ген может мигрировать оттуда, где он есть, туда, где его нет. Таким образом сохранится изменчивость. Учитывая это, Райт сделал вывод, что быстрее развитие будет происходить у тех видов, которые разделены на многочисленные различные по величине популяции. При этом между ними возможна и некоторая миграция. Райт был согласен с тем, что естественный отбор играет весьма существенную роль. Однако вместе с этим результатом эволюции является дрейф генов. Он определяет продолжительные изменения внутри вида. Кроме этого, Райт полагал, что множество отличительных признаков, возникших посредством дрейфа, были безразличны, а в ряде случаев даже вредны для жизнеспособности организмов.